

Species names in all available languages
| Language | Common name |
|---|---|
| Bulgarian | Върбов дрозд |
| Catalan | griveta canyella |
| Croatian | kestenjasti drozd |
| Czech | drozd hnědavý |
| Danish | Rustrygget Skovdrossel |
| Dutch | Veery |
| English | Veery |
| English (United States) | Veery |
| French | Grive fauve |
| French (France) | Grive fauve |
| German | Weidenmusendrossel |
| Greek | Καστανή Θαμνότσιχλα |
| Haitian Creole (Haiti) | Griv pal |
| Hebrew | קיכלי חלוד-גב |
| Hungarian | Vörhenyes fülemülerigó |
| Icelandic | Húmþröstur |
| Japanese | ビリーチャツグミ |
| Lithuanian | Rudagalvis trumpasnapis strazdas |
| Norwegian | viriskogtrost |
| Polish | drozdek brunatny |
| Portuguese (Brazil) | sabiazinho-norte-americano |
| Portuguese (Portugal) | Tordo-fulvo |
| Romanian | Sturz pitic roșcat |
| Russian | Бурый соловьиный дрозд |
| Serbian | Viri drozdić |
| Slovak | drozd hnedkavý |
| Slovenian | Obvodni cikovtnik |
| Spanish | Zorzalito Rojizo |
| Spanish (Argentina) | Zorzalito Colorado |
| Spanish (Chile) | Zorzalito dorsirrojo |
| Spanish (Costa Rica) | Zorzal Dorsirrojizo |
| Spanish (Cuba) | Tordo colorado |
| Spanish (Dominican Republic) | Zorzal Migratorio Colorado |
| Spanish (Honduras) | Zorzalito Rojizo |
| Spanish (Mexico) | Zorzal Canelo |
| Spanish (Panama) | Zorzal Dorsirrojizo |
| Spanish (Paraguay) | Zorzal tropical |
| Spanish (Peru) | Zorzal de Dorso Rojizo |
| Spanish (Puerto Rico) | Zorzalito Rojizo |
| Spanish (Spain) | Zorzalito rojizo |
| Spanish (Venezuela) | Paraulata Cachetona |
| Swedish | rostskogstrast |
| Turkish | Esmer Bülbül Ardıcı |
| Ukrainian | Дрізд-короткодзьоб бурий |
Catharus fuscescens (Stephens, 1817)
Definitions
- CATHARUS
- fuscescens
The Key to Scientific Names
Legend Overview
Veery Catharus fuscescens Scientific name definitions
Version: 1.0 — Published March 4, 2020
Text last updated August 2, 2017
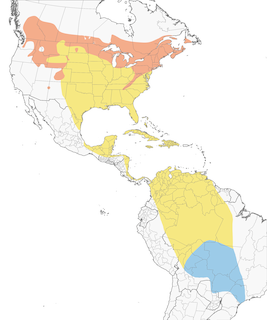
Distribution
Introduction
Breeding Range
Figure 1. Breeds from interior British Columbia and central Alberta, east to southwestern and east-central Ontario (Godfrey 1986, Cadman et al. 2007a), southern Quebec (north to 49°N in west and to just over 50°N in east, and the Anticosti and Magdalen Islands; Falardeau 1996d), the Maritime Provinces (Erskine 1992a) and southwestern Newfoundland south to northern New Jersey, southern Pennsylvania (Wilson et al. 2012), northern Delaware (Hess et al. 2000b), northeastern Maryland, northern half of Ohio (especially northeastern Ohio; Rodewald et al. 2016), northern Indiana, central Minnesota and northern North Dakota (Stewart 1975b). Also breeds in eastern U.S. at higher elevations of the Appalachians from western Maryland south to northern Georgia; e.g., in Tennessee, in far eastern mountains (Great Smokies, Unakas, Blue Ridge) above 900 m, as well as in the Cumberland mountains (Nicholson 1997b). In the western U.S., breeds in Black Hills of western South Dakota (South Dakota Ornithologists' Union 1991), in central and western Montana, Idaho, northern Utah, southern Wyoming, north-central Colorado (localized in mountains at 1,950 to 3,250 m; Wickersham 2016), northeastern Oregon (including Ochoco Mountains, Gilligan et al. 1994), eastern and northern Washington, and along the eastern foothills of the Cascades in Washington (Smith et al. 1997). A disjunct population breeds in some years in vicinity of Little Colorado River in east-central Arizona (Phillips et al. 1964a). Using mitochondrial cytochrome b sequences, Veery showed no sign of recent population expansion between eastern and western populations suggesting the breeding distribution of this species did not change significantly following the last glacial maximum (Topp et al. 2013).
Overwintering Range
Overwintering range encompasses a large area stretching from the southern Orinoco River basin, eastward to Guyana, Suriname, and French Guiana, south to southern Sao Paulo, Mato Grosso and Rondônia States Brazil, and into Bolivia (Remsen 2001, Heckscher et al. 2011, Heckscher et al. 2015). A few records exist from northeastern Paraguay and there is a record from Santa Catarina, Brazil (Kaminski 2011, Pagano et al. 2013). However, these latter records may represent individuals that over-shot their Nearctic–Neotropical or intratropical migrations (Pagano et al. 2013). Recent tracking data from geolocators and miniaturized GPS devices deployed in Delaware and British Columbia, breeding sites show most individuals have two different overwinter areas (Heckscher et al. 2011, Heckscher et al. 2015, Hobson and Kardynal 2015). The species first migrates to a region south of the Amazon River to settles primarily in south-central Brazil on the Brazilian Shield geologic formation. These initial overwinter sites are concentrated in the Brazilian states of Amazonas, Mato Grosso, Para, and Rondônia (Remsen 2001, Heckscher et al. 2015). Veeries then undertake an intratropical migration north, west, and southwest of the Brazilian Shield to a second overwinter site usually in mid-February (Heckscher et al. 2015). Mean arrival of birds at second sites, tracked via geolocators from a Delaware breeding site, was 12 February (range: 5 January–4 March; Heckscher et al. 2015). Second sites are more widespread with most birds moving north of the Amazon River to the Guyana Shield in northern Brazil and southern Venezuela with some settling in Guyana and Suriname. Some birds move south into Bolivia. The two sites together make up the entire overwintering range which encompasses the period between 8 Dec (end of Nearctic–Neotropical migration) and 7 April (onset of Nearctic–Neotropical migration; Heckscher et al. 2011). Of 51 individuals tracked from Delaware only one did not move to a second site and that bird is the only one to date that initially settled north of the Amazon River (CMH). Some Veery “first” sites are in southeastern Brazil in the Atlantic Forest (Tyler 1949b, Remsen 2001) but it is unknown whether these birds also move to second sites. The origin of Atlantic Forest birds is not known considering birds from Delaware and British Columbia – breeding populations nearly 4,000 km apart—have all settled in the Amazon and Orinoco basins. From Delaware geolocator tracking study: mean latitude of first sites 10.4°S ± 3.6 (range 2.3–16.8), mean longitude of first sites 56.4°W ± 4.3 (range 50.8–66.0); mean latitude of second sites 1.6°N ± 5.9 (range 2.9°N–17.5°S), mean longitude of second sites 60.1°W ± 4.2 (range 50.3–68.7) (Heckscher et al. 2015). Duration at initial overwinter sites ranged from 36–121 d; duration at second sites ranged from 38–97 d (Heckscher et al. 2015).
Note earlier descriptions of the species’ overwintering range included northern Colombia and other areas of South America (e.g., American Ornithologists' Union 1983) but these descriptions were based on transient individuals rather than settled birds (Remsen 2001). It was suggested that Veery overwintering range should only include 2 concentrated areas in southern Brazil and the Atlantic Forest (Remsen 2001), but that assessment was prior to geolocator studies that showed Veery moves to a second overwinter site after first settling in Remsen’s proposed regions of southern Brazil (Heckscher et al. 2011).
Extralimital Records
Accidental in Britain and Sweden (Alström and Colston 1991).
- Year-round
- Migration
- Breeding
- Non-Breeding

eBird Status and Trends

















