

Species names in all available languages
| Language | Common name |
|---|---|
| Bulgarian | Очилат дрозд |
| Catalan | griveta costanera |
| Croatian | patuljasti drozd |
| Czech | drozd západní |
| Danish | Olivenskovdrossel |
| Dutch | Dwerglijster |
| English | Swainson's Thrush |
| English (United States) | Swainson's Thrush |
| Finnish | korpirastas |
| French | Grive à dos olive |
| French (France) | Grive à dos olive |
| French (Haiti) | Grive de Olive |
| German | Zwergmusendrossel |
| Greek | Κοινή Θαμνότσιχλα |
| Hebrew | קיכלי זיתני |
| Hungarian | Flótázó fülemülerigó |
| Icelandic | Moldþröstur |
| Italian | Tordo di Swainson |
| Japanese | オリーブチャツグミ |
| Lithuanian | Svaisono strazdas |
| Norwegian | brunkinnskogtrost |
| Polish | drozdek okularowy |
| Portuguese (Brazil) | sabiazinho-de-óculos |
| Portuguese (Portugal) | Tordo-dos-pântanos |
| Romanian | Sturz pitic melodios |
| Russian | Дрозд Свэнсона |
| Serbian | Žutogrli drozdić |
| Slovak | drozd olivový |
| Slovenian | Smrekov cikovtnik |
| Spanish | Zorzalito de Swainson |
| Spanish (Argentina) | Zorzalito Boreal |
| Spanish (Chile) | Zorzalito de Swainson |
| Spanish (Costa Rica) | Zorzal de Swainson |
| Spanish (Cuba) | Tordo de espalda olivada |
| Spanish (Dominican Republic) | Zorzal de Swainson |
| Spanish (Ecuador) | Zorzal de Swainson |
| Spanish (Honduras) | Zorzalito de Anteojos |
| Spanish (Mexico) | Zorzal de Anteojos |
| Spanish (Panama) | Zorzal de Swainson |
| Spanish (Peru) | Zorzal de Swainson |
| Spanish (Puerto Rico) | Zorzalito de Swainson |
| Spanish (Spain) | Zorzalito de Swainson |
| Spanish (Venezuela) | Paraulata Lomiaceituna |
| Swedish | beigekindad skogstrast |
| Turkish | Bülbül Ardıcı |
| Ukrainian | Дрізд-короткодзьоб Cвенсона |
Catharus ustulatus (Nuttall, 1840)
Definitions
- CATHARUS
- ustulata / ustulatus
The Key to Scientific Names
Legend Overview
Swainson's Thrush Catharus ustulatus Scientific name definitions
Version: 1.0 — Published March 4, 2020
Text last updated January 1, 2000
Breeding
Phenology
Pair Formation
Arrival varies by location and elevation, with males preceding females: mid- to late Apr in coastal California (147); mid-May at a midelevation Idaho site (DEM) and in se. Alaska (148); and about mid-May in Massachusetts (56). Only Nearctic-Neotropical migratory thrush species whose second-year and older males may begin releasing sperm into cloaca spontaneously and continuously prior to arrival on some breeding grounds (documented 800 km south of nesting range), possibly as mechanism for preparation for appropriately timed mating (119).
Nest-Building
Figure 3 . Mean date of nests observed under construction (not backdated from hatch) 18 Jun ± 7.5 d SD in Idaho (range 9 Jun–5 Jul, n = 21; DEM, J. Bednarz, J. Hovis, and C. Davis unpubl.); individuals with nesting material observed 3–16 May in Monterey, CA (83). Limited data indicate nest built at least 2–3 wk after arrival at breeding sites: e.g., approximately 20 d in Idaho (DEM) and Maine (10).
First/Only Brood Per Season
Across the range, eggs laid mid-Apr–late Jun, although later dates recorded for what are presumed to be renests from early failures. Mean dates (± SD) of eggs in nests (generally the incubation period, not necessarily first day of incubation) from combined sources (Western Foundation of Vertebrate Zoology [WFVZ], Cornell nest record program [CNRP], and 96: 3 Jun ± 16 d (range 18 Apr–25 Jul, n = 345) in California; 18 Jun ± 14 d (range 29 May–12 Jul, n = 20) in Alaska; 21 Jun ± 11 d (range 28 May–25 Jul, n = 83) in the Pacific Northwest; 20 Jun ± 9 d (range 21 May–14 Jul, n = 90) in the Northeast, and 23 Jun ± 9 d (range 17 May–19 Jul, n = 87) in Inland North-west. In British Columbia, 51% of 328 nests had eggs present 14–30 Jun (42). Mean date of first egg 10 Jun ± 2.4 d SE for Golden Gate National Recreation Area, CA (range 11 May–Jul 6, n = 33; 149); mean date of nest initiation 27 Jun ± 7 d (range 6 Jun–5 Jul, n = 25) in Green Mtns., VT (K. McFarland pers. comm.).
Young observed in nests, from combined sources: 10–26 Jun in Alaska (n = 3), 10 Jun–6 Jul in Northeast (n = 10), 22 Jun–24 Jul in e.-central U.S. (n = 8), 17 Jun–29 Jul in Inland Northwest (n = 56), and 24 Jun–1 Aug in Pacific Northwest (n = 17). (Forward- and backdating from incubation and fledging dates could extend this period for some locations.) Of British Columbia nests, 52% had young between 29 Jun and 15 Jul (42). Fledglings 23 Jun–3 Aug in Monterey, CA (83); mean fledge date for Idaho study 20 Jul ± 9 d (range 8 Jul–13 Aug, n = 42; 96).
Second Brood Per Season
Generally 1 brood/season but may renest in some localities if first clutch is lost early (2 egg sets reportedly collected from same pair in California; WFVZ). A slight second peak in number of broods 3 wk after first peak suggested as evidence of double-brooding in British Columbia (42). One female at a low-elevation coastal site in British Columbia successfully fledged young and then laid a second clutch (which failed immediately), giving the only direct evidence of an attempt at a second brood (J. Smith pers. comm.). At higher elevations, snow pack limits breeding season to 6–9 wk, allowing insufficient time for a second brood.
Nest Site
Selection
No information.
Microhabitat/Site Characteristics
Most frequently in understory, particularly in thickets of deciduous shrubs or conifer saplings; less frequently but consistently found >3 m high on top of a horizontal branch away from the bole of a larger-diameter tree. Of 981 nests from combined sources (42, 96, WFVZ, CNRP, K. McFarland pers. comm.): height range 0–24 m; 60% between 1.0 and 2.4 m. Higher placement more typical of conifer forest sites. More than 90% of nests (n = 386) in California and Pacific Northwest <3 m, while 53% (n = 109) of Inland Northwest nests ≥3 m; 70% (n = 113) of nests in ne. North America between 1 and 2.5 m. Mean height in Alaska taiga 1.8 m ± 1.0 SD (n = 121; 9). Nests of isolated population in Allegany Hills, NY, characteristic of high nests, averaging 5.8 m high, 3.6 m from bole of large eastern hemlock (Tsuga canadensis) with limited understory (150). In Idaho, 29% nests of this type (96).
Wide variety of plant species used as substrate; most frequently reported (sources listed above except 42) include willow (17%), fir (15%), spruce (14%), blackberry (11%), and unidentified bush (11%). Also used other deciduous trees, including alder, aspen, birch (Betula sp.), maple (Acer sp.), and oak; deciduous shrubs, such as briers, gooseberry (Ribes sp.), rose (Rosa sp.), sumac, and vinemaple (Acer sp.); and coniferous trees, including cedar (Thuja sp.), hemlock, and lodgepole pine (Pinus contorta). Unusual nests observed on a piece of driftwood, on the ground in grass, in a rootwad, and on a rock base. One nest found in small snag within dense thicket of pole-sized trees. Active nest (full clutch) reported from Vermont built atop artificial nest with quail eggs (K. McFarland pers. comm.). Stumps and posts (6 records) used when enhanced by vine cover.
In mixed-conifer forest in Idaho, nests placed nearer water in areas with denser shrubs and saplings, more trees 8–23 cm diameter, more large trees (>38 cm dbh), and greater canopy cover (85% ± 13.5 SD) than random sites (96). Nest sites had more grand fir trees (Abies grandis), and this substrate supported 59% of 117 nests. No attributes measured were significantly associated with nest fate. Similar characteristics reported from Vermont: 86% ± 11.35 SD canopy cover, higher density of small-, medium-, and large-diameter balsam fir (Abies balsamea) than other tree species, and 72% of 25 nests placed in fir (K. McFarland pers. comm.). Nest-plot attributes selected were large birch trees and large and small conifer trees. Microsite attributes for nests in western coniferous and eastern mixed coniferous-deciduous forests compared in Appendix 1 .
From riparian sites in central coastal California, nest sites associated most strongly with canopy cover; red alder (Alnus rubra) density; larger tree density; and fern, salmonberry (Rubus sp.), and hedge nettle (Stachys sp.) cover. Negatively associated with artichoke thistle (Cynara cardunculus) cover, grass/sedge cover, and logs (S. Small, G. R. Geupel, N. Nur, A. L. Holmes, and T. Gardali unpubl.). More than 30% of nests placed in California blackberry (Rubus ursinus); hedge nettle, salmonberry, and lady fern (Athyrium filix-femina) also selected. Nest success positively influenced by total canopy cover, red alder tree density, western swordfern (Polystichum munitum) cover, and nest concealment from below; willow-shrub cover, snag density, forb cover, and shrub species richness had negative associations with success (149; S. Small, G. R. Geupel, N. Nur, A. L. Holmes, and T. Gardali unpubl.).
Nest
Construction Process
Constructed in about 4 d by female (1, 3). No information on methods or time of day. Process of shaping nest cup by snuggling low into cup and thrusting with breast and feet inferred from other thrushes (e.g., Wood Thrush, 35).
Structure And Composition Matter
Outer layer of nest composed of grasses, plant stems, moss, small pieces of bark, small twigs; inner lining of skeletonized leaves, rootlets, lichens, or moss (10, 78, 151). Mud often mentioned in literature, but not observed in Idaho or Vermont nests (K. McFarland pers. comm., DEM), and grass and moss predominated in nests from British Columbia (42). External appearance varies. Described at times as conspicuous (10), but in general, nests difficult to find, as evidenced by few nest discoveries relative to other species in Breeding Bird Atlas efforts. Characteristic nests in Idaho are small to bulky, light brown–colored grassy cups, and new nests identified by pieces of newly woven grass protruding out and hanging down from nest cup (DEM). In coastal Pacific Northwest, green moss replaces grass and has a draped effect. Other nests predominantly of small twigs (1). Rhizomorphs of horsehair fungus (Marasmius androsaceus) discovered as common nest-lining in spruce-fir forests in New England (152); these authors suggested previous descriptions of “rootlets” in nest-linings may be this material. Fungus known to have antibiotic capabilities (i.e., could function in deterring nest pathogens).
Dimensions
Outside diameter 9.4–15.4 cm (10.6 ± 1.32 SD), height 3.8–10.5 cm (8.2 ± 1.7 SD); inside diameter 5.0–8.5 cm (6.0 ± 0.85 SD), depth 2.5–6.0 cm (3.8 ± 0.98 SD) from 19 nests in Green Mtns., VT (K. McFarland et al. unpubl.). Some nests in Idaho outside these reported ranges: outside diameter 16.5 cm, depth 1.75 cm (DEM, J. Bednarz, J. Hovis, and C. Davis unpubl.).
Microclimate
Usually placed in shady location. No information on temperature or moisture at nests.
Maintenance Or Reuse Of Nests, Alternate Nests
New nest built each season, and also if renest attempted after failure within a season (DEM). Nest structures occasionally persist through winter in Idaho, but no reuse documented.
Nonbreeding Nests
Not known to be used. May build and then abandon a nest before eggs laid, possibly due to disturbance (2 nests built near frequently used hiking trails abandoned; 96).
Eggs
Shape
Ovate, varying to short and long ovate (1).
Size
Length 20.2–25.7 mm (mean 23.19 mm, calculated from subspecies means); breadth 15.27–18.07 mm (mean of subspecies means 16.66 mm, n = 238 eggs, 66 clutches in WFVZ collection). C. u. swainsoni and ustulatus similar; clarescens (now swainsoni) on average 1.5 mm longer.
Mass
No information.
Color
Blue to greenish blue with reddish or brown speckles (151). Density and size of spots variable, from uniformly covering egg to concentration toward larger end (10, 1). Brown-on-blue color combination common among species that nest in fork of bush or tree and that often have nests with dark linings; egg placement on dark lining could reduce visibility to predators (153).
Surface Texture
Smooth.
Eggshell Thickness
Shell thickness not available. Empty shell weight: 0.140–0.218 g, mean of sub-species means 0.182 g (WFVZ).
Clutch Size
One to 5 eggs; 4 eggs most common (61% of 911 nests; 42, 96, CNRP, WFVZ).
Egg-Laying
Begins after nest completion. Number of days not well quantified; within 5 d at nests in Idaho (DEM). One egg/d laid in morning (10). No information on parental behavior during this period or egg replacement. No evidence of intraspecific egg-dumping.
Incubation
Onset Of Broodiness And Incubation In Relation To Laying
Female begins incubating with last egg laid, but young still hatch asynchronously (10).
Incubation Patch
Of Swainson's Thrushes captured on breeding sites, females had visible brood patch, no brood patch observed on males in the hand (J. Bednarz pers. comm.).
Incubation Period
Ranges 10–14 d from last egg laid (42, 96). Young found on days 10, 11, 12, and 13 from onset of incubation of Maine nests (10).
Parental Behavior
Only female incubates (10). Female cautious while approaching nest and generally will not go if she detects an observer or is emitting a Whit Call. Approaches nest from near ground, moves up to nest limb, then quickly goes to nest; departs by flying down and away. Individuals vary in response to humans by flushing or sitting tight when incubating, but tend to sit very tight when eggs are hatching or nestlings are 1–2 d old (DEM). Female generally does not vocalize while on nest (D. Masingale pers. comm.). Male delivers food to female at nest (DEM), although frequency and extent to which female relies on male for food not reported. Male sings nearby throughout incubation period (10, DEM). No information on rhythm or duration of female attentive periods.
Hardiness Of Eggs Against Temperature Stress; Effect Of Egg Neglect
No information.
Hatching
Very little information. From 2 4-egg nests, 7 of 8 young were newly hatched in mid- to late morning (09:30–11:55; 10). In each nest, first 2 young hatched together, suggesting partial synchrony, but 3 of remaining 4 were approximately 24 h apart (48 h between first and last hatch). Egg-shells removed shortly after hatch, presumably by female, but no documentation.
Young Birds
Condition At Hatching
From Stanwood (10). Altricial, partially covered with natal down, eyes closed. Body length approximately 4 cm, mass 3.9 g, wing length 0.3 cm. Natal down dark, burnt amber, 1 cm long; beak, legs, and feet tinged burnt orange. Young lay limp in nest cup, ≥2 curled together “vibrating as one.”
Growth And Development
Few data. Observations of nests in Maine (10) found eyes beginning to open 2 d after hatching; feather tracts obvious by end of day 3; quills and pin-feathers evident by day 5; pin-feathers beginning to show along feather tracts; heads speckled by day 7; well feathered by end of day 10. Body length more than doubled in 10 d, increasing to 9.5 cm; wing length increased to 5.6 cm, mass increased 6-fold to >25 g. Moving in nest, panting, and soft vocalizations in response to food by day 4; preening by day 7; frequent movement in and on nest, including wing flaps, by end of day 10. Degree to which nestlings beg for food variable; noticeably audible in Maine and Vermont (10, K. McFarland pers. comm.), but not recorded in Idaho (J. Bednarz pers. comm., DEM).
Parental Care
Brooding
Female broods closely for first 3–4 d, frequently grooming young (10). At approach of human observer, sits in low incubating position, entirely still but alert, and may remain even when observer brushes past nest (DEM). As young develop, female is off nest more frequently and for longer periods of time (10); no quantitative information on duration, rhythm, or relationship to weather. Will still brood young 10–11 d old (DEM).
Feeding
Both male and female share feeding responsibilities from hatch (10), although proportion of effort not quantified. Initially feed regurgitated food as well as fresh insects. In 3 h, adults made 20 visits to a brood of 4 young 3–6 d old (10). Daily feeding frequency (n = 2 nests, age unknown) calculated at 72 feedings/adult/d, based on each nestling being fed 48 times in 14 h, equating to 144 feedings to a brood of 3 (6). Animal food, primarily caterpillars, beetles, moths, flies, and other bugs, made up higher percent of nestling diet (93%) than adult diet (63%); fruits (raspberries and blackberries) relatively small proportion.
Nest Sanitation
Female appears to eat or remove parasites from nest and bodies of young by grooming with bill. Both adults consume fecal sacs in early nestling period; begin to remove them from nest about 9 d after hatching (10).
Cooperative Breeding
Not known to occur.
Brood Parasitism by Other Species
Identity Of The Parasitic Species
Brown-headed Cowbird only documented brood parasite for this species.
Frequency Of Occurrence, Season Or Geographic Variation
No Brown-headed Cowbird eggs in 526 nests (348 of these from California) in the WFVZ collection spanning 1867–1970, and only 6 inci-dences of cowbird eggs from 98 records of CNRP dating from 1894 to 1996. Current parasitism rates vary geographically and may reflect the changing distribution of cowbirds in some parts of U.S. Declines in Swainson's Thrush populations along central and s. California coast (see Distribution: historical changes, above) have been attributed in part to cowbird brood parasitism (45; L. Kiff, S. Laymon pers. comm.), although 0 of 39 nests monitored in 1995–1996 at Point Reyes National Seashore, CA, contained cowbird eggs (154), and only 2 of 65 nests (3%) were parasitized across central Californian riparian sites in 1997 (S. Small, G. R. Geupel, N. Nur, A. L. Holmes, and T. Gardali unpubl.). Rates were slightly higher (8%; n = 38) at 2 drainages in Golden Gate National Recreation Area (149). Similar rate (8%) reported from a large sample in British Columbia (n = 395), with little difference between inland and coastal regions (42). Of 110 nests with known outcome monitored in w.-central Idaho from 1994 to 1998, only 1 was parasitized by cowbirds (96). No parasitism in a small sample (n = 7) of nests in se. Alaska (and in 42 nests of 3 other species of open-cup nesters in same area; 148), and no parasitism documented from 25 nests in New York and Vermont forests during 1996–1997 (K. McFarland pers. comm.).
Timing Of Laying In Relation To Host'S Laying
Brown-headed Cowbird egg(s) observed laid after Swainson's Thrush egg 1, 2, and 3 (WFVZ, CNRP).
Response To Parasitic Mother, Eggs, Or Nestlings
No reported interaction with Brown-headed Cowbird female. Limited data provide no pattern of response, which included continued incubation of all eggs but successfully raising only 1 cowbird (n = 3); abandonment after cowbird egg laid (n = 3); and removing the egg and continuing incubation and brooding (n = 1; CNRP, J. Smith pers. comm.). Up to 3 cowbird eggs reported from a single nest.
Effects Of Parasitism On Host
Despite egg loss from individual nests, data do not suggest that parasitism is suppressing populations through decreased reproductive success, even in California; declines in Sierra Nevada preceded cowbird presence there (R. Stefani pers. comm.). Across range, more nests fail from causes other than parasitism: 14% of 36 nests with known outcome in CNRP database failed due to Brown-headed Cowbirds, while 42% failed from predation and 31% successfully fledged at least 1 Swainson's Thrush. Failure rates approached 60% in Idaho, none due to parasitism.
Success Of Parasite With This Host
Limited data suggest that Brown-headed Cowbirds have low success with this species, based on relatively low rates of parasitism and high rates of failure of parasitized (and all) nests (see above).
Fledgling Stage
Few data. Leaves nest after 10–14 d; fledged at 10 d at a nest where young were removed and replaced daily and parents agitated, 12–14 d at undisturbed nests (10, 42, 96). Active in nest prior to fledging, with frequent stretching and preening, hopping from nest cup to rim (10). Fledging event not reported; role of parents in triggering departure from nest (vocal encouragement, food enticement, withholding food) unreported, although at 1 frequently disturbed nest, parents called repeatedly to young after they were replaced in nest, and nest was empty 1 h later (10). Departs nest in Juvenile plumage; capable of standing, perching, and short, fluttery flight. Fledglings remain close to nest tree for 1–2 d, begin to wander subsequently (DEM). One fledgling observed 300 m from nest 5 d postfledging at a site in Idaho, being fed by a banded parent (J. Hovis pers. comm.). Duration of dependence on and association with parents unknown; 21–31 d for Wood Thrush (35).
Immature Stage
No information.
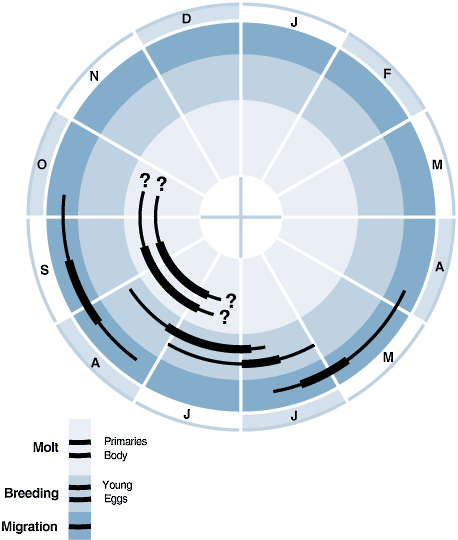
Annual cycle of Swainson’s Thrush breeding, molt, and migration for interior nw. North America (central Idaho to s. British Columbia). Geographic variation is described in the text. Thick lines show peak activity; thin lines, off-peak.

San Diego Co., CA. 31 May 1920. Ruler is in cm.; photographer Rene Corado

Ventura Co., CA. 16 May 1919. Ruler is in cm.; photographer Rene Corado

















