












Surf Scoter Melanitta perspicillata Scientific name definitions
- LC Least Concern
- Names (38)
- Monotypic
Eric M. Anderson, Rian D. Dickson, Erika K. Lok, Eric C. Palm, Jean-Pierre L. Savard, Daniel Bordage, and Austin Reed
Version: 1.0 — Published March 4, 2020
Text last updated May 7, 2015
Text last updated May 7, 2015
Sign in to see your badges
Species names in all available languages
| Language | Common name |
|---|---|
| Asturian | Zapateru caretu |
| Basque | Ahatebeltz kopetazuria |
| Bulgarian | Черна потапница |
| Catalan | ànec negre frontblanc |
| Croatian | lisasta patka |
| Czech | turpan pestrozobý |
| Danish | Brilleand |
| Dutch | Brilzee-eend |
| English | Surf Scoter |
| English (United States) | Surf Scoter |
| Faroese | Nakkahvít ont |
| Finnish | pilkkaniska |
| French | Macreuse à front blanc |
| French (France) | Macreuse à front blanc |
| Galician | Mourelo careto |
| German | Brillenente |
| Greek | Αμερικανική Μαυρόπαπια |
| Hebrew | קטיפנית משברים |
| Hungarian | Pápaszemes réce |
| Icelandic | Krákönd |
| Japanese | アラナミキンクロ |
| Korean | 흰이마검둥오리 |
| Latvian | Bangu pīle |
| Lithuanian | Baltakaktė nuodegulė |
| Norwegian | brilleand |
| Polish | uhla pstrodzioba |
| Portuguese (Portugal) | Negrola-de-lunetas |
| Romanian | Rață neagră cu ochelari |
| Russian | Пестроносый турпан |
| Serbian | Lisasti turpan |
| Slovak | turpan okuliarnatý |
| Slovenian | Očalarka |
| Spanish | Negrón Careto |
| Spanish (Mexico) | Negreta Nuca Blanca |
| Spanish (Spain) | Negrón careto |
| Swedish | vitnackad svärta |
| Turkish | Yamalı Ördek |
| Ukrainian | Турпан білолобий |
Melanitta perspicillata (Linnaeus, 1758)
PROTONYM:
Anas perspicillata
Linnaeus, 1758. Systema Naturæ per Regna Tria Naturæ, Secundum Classes, Ordines, Genera, Species, cum Characteribus, Differentiis, Synonymis, Locis. Tomus I. Editio decima, reformata 1, p.125.
TYPE LOCALITY:
Hudson Bay, ex Edwards.
SOURCE:
Avibase, 2023
Definitions
- MELANITTA
- perspicillata / perspicillatum / perspicillatus
The Key to Scientific Names
Legend Overview
UPPERCASE: current genus
Uppercase first letter: generic synonym
● and ● See: generic homonyms
lowercase: species and subspecies
●: early names, variants, misspellings
‡: extinct
†: type species
Gr.: ancient Greek
L.: Latin
<: derived from
syn: synonym of
/: separates historical and modern geographic names
ex: based on
TL: type locality
OD: original diagnosis (genus) or original description (species)
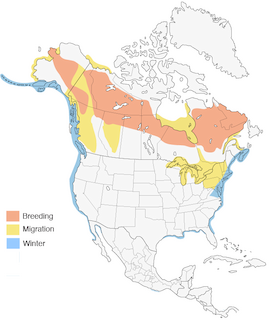
- Year-round
- Migration
- Breeding
- Non-Breeding
Figure 1. Distribution of the Surf Scoter.















