

Species names in all available languages
| Language | Common name |
|---|---|
| Albanian | Gjelaci |
| Arabic | دريجة شبه مكففة |
| Asturian | Mazaricu semipalmeñu |
| Bulgarian | Пясъчен брегобегач |
| Catalan | territ semipalmat |
| Croatian | žalar pješčar |
| Czech | jespák srostloprstý |
| Danish | Tyknæbbet Dværgryle |
| Dutch | Grijze Strandloper |
| English | Semipalmated Sandpiper |
| English (United States) | Semipalmated Sandpiper |
| Faroese | Fitjugrælingur |
| Finnish | kanadansirri |
| French | Bécasseau semipalmé |
| French (France) | Bécasseau semipalmé |
| Galician | Pilro semipalmado |
| German | Sandstrandläufer |
| Greek | Καναδέζικη Νανοσκαλίδρα |
| Haitian Creole (Haiti) | Ti Bekasin janm nwa |
| Hebrew | חופית אמריקנית |
| Hungarian | Kis partfutó |
| Icelandic | Fitjatíta |
| Italian | Gambecchio semipalmato |
| Japanese | ヒレアシトウネン |
| Lithuanian | Tundrinis bėgikas |
| Norwegian | sandsnipe |
| Polish | biegus tundrowy |
| Portuguese (Brazil) | maçarico-rasteirinho |
| Portuguese (Portugal) | Pilrito-rasteirinho |
| Romanian | Fugaci semipalmat |
| Russian | Малый песочник |
| Serbian | Poluopnasta sprutka |
| Slovak | pobrežník piesočný |
| Slovenian | Krpljasti prodnik |
| Spanish | Correlimos Semipalmeado |
| Spanish (Argentina) | Playerito Enano |
| Spanish (Chile) | Playero semipalmado |
| Spanish (Costa Rica) | Correlimos Semipalmado |
| Spanish (Cuba) | Zarapico semipalmeado |
| Spanish (Dominican Republic) | Playerito Semipalmado |
| Spanish (Ecuador) | Playero Semipalmeado |
| Spanish (Honduras) | Playerito Semipalmado |
| Spanish (Mexico) | Playero Semipalmeado |
| Spanish (Panama) | Playero Semipalmeado |
| Spanish (Paraguay) | Playerito escudado |
| Spanish (Peru) | Playerito Semipalmado |
| Spanish (Puerto Rico) | Playero Gracioso |
| Spanish (Spain) | Correlimos semipalmeado |
| Spanish (Uruguay) | Playerito Enano |
| Spanish (Venezuela) | Playerito Semipalmeado |
| Swedish | sandsnäppa |
| Turkish | Perdeli Kumkuşu |
| Ukrainian | Побережник тундровий |
Calidris pusilla (Linnaeus, 1766)
Definitions
- CALIDRIS
- calidris
- pusilla
The Key to Scientific Names
Legend Overview
Semipalmated Sandpiper Calidris pusilla Scientific name definitions
Version: 1.0 — Published March 4, 2020
Text last updated February 24, 2010
Breeding
Phenology
Pair Formation
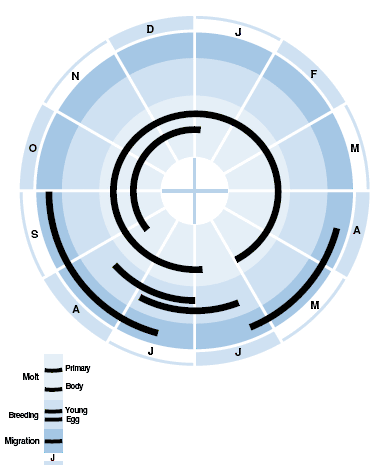
See Figure 3. Arrive on breeding grounds in Manitoba and Alaska last week of May to early June (Dixon 1943, Littlefield and Pakulak 1969, Ashkenazie and Safriel 1979, Jehl 2006). Males normally precede females by less than a week, and set up territories almost immediately (CLG-T). Most pairs form shortly afterward (Ashkenazie and Safriel 1979), although some new birds appear late and pair then, if possible (CLG-T).
Nest Building
In both Alaska and n. Manitoba, birds appear on breeding territories first days of June and nest 7-10 June (Ashkenazie and Safriel 1979; Gratto and Cooke 1987; Jehl 2004a). Pairs engage for several days in nest-scraping, where males create numerous scrapes.
First/Only Brood Per Season
First egg laid 4 to 6 d after pair formation (Ashkenazie and Safriel 1979). Timing of egg-laying appears dependent on availability of invertebrate prey, as laying can be delayed for several weeks in inclement weather. In two years of severe weather at La Pérouse Bay, Manitoba, approximately half of paired females did not lay clutches (Gratto-Trevor 1991).
First egg in Manitoba, Alaska, and Victoria I., NWT: 2 to 20 June (Brandt 1942, Dixon 1943, Parmelee et al. 1967, Ashkenazie and Safriel 1979, CLG-T). Peak nest initiation usually third week in June (Parmelee et al. 1967, Ashkenazie and Safriel 1979, Gratto and Cooke 1987). Although snow melt averages almost a month earlier at La Pérouse Bay, Manitoba, than Barrow, AK (18 May versus 12–17 June), laying occurs at the same time in both areas. Peak insect hatch also occurs at the same time in both areas (McClure 1943a). Late arrivals and renesting attempts may extend nest initiation period to early July. Clutches initiated later than 1 July often are deserted late in incubation (CLG-T, Holmes and Pitelka 1968).
In both Alaska and n. Manitoba, earliest hatch in last few days of June (Brandt 1942, Dixon 1943. In both Alaska and n. Manitoba, earliest hatch in last few days of June Brandt 1942, CLG-T), after 20 days of incubation. Peak hatch in both areas normally second week in July (Norton 1973, Gratto and Cooke 1987). Most chicks fledge in late July, at 16–19 d of age (Safriel 1975, Gratto-Trevor 1991). Although widely separated geographically, breeding chronology, from pair formation to fledging, is similar in both regions.
Second Brood Per Season
Rarely observed to lay or incubate two clutches in one season, unless first clutch is predated or deserted early in breeding season (CLG-T). Renests can be laid as soon as 5 d after clutch is lost (Safriel 1971). In Churchill, Manitoba, an experienced pair lost a 4-egg clutch to flooding on 14 June and produced a full replacement clutch 8 d later (Jehl 2007). Because Manitoba nests due to hatch after 24 July are often deserted, less than two weeks are available from completion of first clutch to initiation of renest (CLG-T).
Nest Site
Selection Process
Males establish territory, normally the same as previous year, before females arrive (Gratto et al. 1985). Males may make scrapes before pair formation, as well as after. Females select scrape.
Microhabitat
No information.
Site Characteristics
In river deltas in Manitoba and Alaska, males select nest locations near ponds as well as along the sides or tops of small hummocks or ridges. These could be located under, or adjacent to, small willow (Salix) bush or sweetgale (Myrica gale), dwarf birch (Betula nana) or in a clump of sedges (Carex spp.) in damp low areas (Gratto-Trevor 1992; Jehl 2006) -- but not on barren sandy flats or rocky ridges (Brandt 1942, Savile 1951, Parmelee et al. 1967). Nest sites generally drier than those of Dunlin and Red-necked Phalaropes, which nest in the same habitat (R. Gill pers. comm., CLG-T).
On islands in e. Hudson Bay, nests in slight depressions in low sedge-tundra; also on higher ridges, among tufts of grass growing in fine gravel (Todd 1963a).
Nest
Construction Process
Nest scraping displays common in prelaying birds, where males make scrapes by pressing breast against ground and rotating body. Female examines scrapes and lines one with small leaves or grass using sideways building display, by sitting in nest cup and throwing lining material over back. Copulation occurs after nest selection. Female continues to add lining material throughout the laying period (Ashkenazie and Safriel 1979).
Structure And Composition Matter
Nest mere depression in ground, lined with dead willow, dwarf birch (Betula spp.) or cranberry leaves (Oxycoccus spp.), and pieces of grass, sedge or moss (Brandt 1942, Savile 1951, Parmelee et al. 1967).
Dimensions
Inside diameter is 5–6 cm, inside depth 4–5 cm (Brandt 1942).
Microclimate
No data.
Maintenance Or Reuse Of Nests, Alternative Nests
Previous year's nest cup commonly reused if young successfully hatched there. Old nest cups often rescraped but not reused. Occasionally use nest cups of other pairs (if other birds not present), or even those of other species such as Dunlin, Red-necked Phalarope, Horned Lark (Eremophila alpestris), and Savannah Sparrow (Passerculus sandwichensis; Gratto et al. 1985). Site of depredated nest not reused by same birds in same year (CLG-T).
One male sandpiper in Nome, Alaska, mated with three different females in three consecutive years and reused the same nest cup each time (Sandercock et al. 2000). In Churchill, Manitoba, one pair used the same nest for four successive years (Jehl 2006). The re-use of old nests allows pairs to nest earlier and with greater success. Birds with high rates of territory, mate and nest-site retention breed earlier and nest as soon as habitat conditions permit. Fledglings of experienced early-nesters survive better than later-fledged conspecifics (Jehl 2006).
Nonbreeding Nests
Nest cup provided by male for brooding chicks each night for first 6–8 days after hatch (Ashkenazie and Safriel 1979).
Eggs
Shape
Ovate pyriform to subpyriform (Bent 1927, Brandt 1942). Because of pyriform shape, with small ends together, 4 eggs of clutch fit together snugly in nest cup.
Size
Eggs of adult pairs in Manitoba: n = 310, length 30.0 ± 0.9 mm, breadth 21.3 ± 0.5 mm; calculated volume (length x breadth2) 13.6 mm3. Eggs at nests of yearling females smaller in length but not breadth (Gratto et al. 1983). Runt eggs, such as described by Manning and Carter (Manning and Carter 1977: 22.2 x 16.1, 42% normal volume) rare; none noted in Manitoba among > 1,800 eggs examined (CLG-T).
Average mass of fresh egg 7.3 g near Barrow, AK, dry weight excluding shell 1.5 g, and 46% fat (MacLean 1969). Mass of fresh egg in Manitoba averaged only 6.3 g (CLG-T). Calcium content of clutch twice that of prelaying female (MacLean 1974). Mass of an egg 22%–25% of average female mass (28.7 g, both Alaska and Manitoba; Ashkenazi and Safriel 1979, Gratto 1983). Therefore, weight of 4–egg clutch = 88%–100% of female's weight.
In eggs collected from 1914 to 1947 in Alaska, mean egg thickness index ± SE = 0.46 ± 0.002 mm (99 eggs of 26 clutches), and from 1951 to 1965, 0.46 ± 0.004 mm (77 eggs of 20 clutches): no difference between periods (Morrison and Kiff 1979).
Color
Color and pattern of eggs varies greatly among clutches, even in a single breeding area (CLG-T). Not known whether patterns are consistent for individual females. Eggs cannot be definitely distinguished from those of Least or Western sandpipers, as measurements, coloring, and markings intergrade with both species (Bent 1927, Gabrielson and Lincoln 1959).
Ground color varies from dull white to pale olive buff and olive buff, occasionally to deep olive buff or isabella. Markings liver brown to chestnut brown in darkest, and hazel to cinnamon brown in lightest. Markings often have spiral tendency, particularly at large end of egg, but many spots round instead of elongate. Underlying markings light quaker drab to quaker drab (Bent 1927, Brandt 1942).
Surface Texture
Smooth and glossy (Harrison 1978a).
Egg Laying
Some energy for egg production acquired during migration (MacLean 1969), but more must be obtained on breeding grounds. First egg can be laid 4–6 d after pairing (Ashkenazie and Safriel 1979), if food available, otherwise laying may be delayed for as much as two weeks, and some paired females may not lay at all in some years (Gratto-Trevor 1991). In Alaska, egg-laying intervals greater than 30 h (0.8 eggs/d) such that this northerly breeding population (64º N) has a slower egg-laying rate (mean 28.6 h ± 7.2) than the more southern breeding population in Manitoba (1 egg/day) (Gratto-Trevor 1992; Sandercock 1998b; Sandercock et al. 1999).
Overall, laying intervals range from < 24 h up to 32 h, such that a 4-egg clutch is laid over 4-5 d. The northern population often produces clutches with less than 4 eggs and birds rarely renest, whereas the more southerly breeding population almost always has 4 eggs and birds renest more often (Gratto-Trevor 1992). This may be due to the earlier (3-4 wk) nesting schedule in Alaska (Sandercock 1998b).
Limited fixed clutch size of 4 eggs may be due to selection on factors acting during egg production or brood-rearing -- regulating offspring number (Sandercock 1997b). Eggs deposited often in morning, evening or night (Ashkenazie and Safriel 1979). Females do not store calcium in medullary bone, but acquire it from insect prey and ingestion of small mammal bones. After laying, females contain less calcium than males or prelaying females, and decalcified areas are often visible in their skulls (MacLean 1974).
Males normally follow females during the laying period, from feeding to nest sites, except for occasional territorial displays and short spells of incubation (Ashkenazi and Safriel 1979).
No evidence for replacement of eggs destroyed during laying. In all instances where an egg is broken before laying of third or fourth egg, broken egg is removed, and final clutch size is only three. About 7% of eggs that fail to hatch remain abandoned in nest cup (Sandercock 1996). Intraspecific egg dumping never observed, and never more than four eggs of this species in almost 600 nests examined (CLG-T).
Incubation
Onset Of Broodiness And Incubation In Relation To Laying
Partial incubation by both sexes during laying, with continuous incubation several hours before laying of last egg (Ashkenazie and Safriel 1979), although nest attendance and incubation may be initiated before laying is complete (Sandercock 1998b).
Incubation Patches
Incubating males and females have two large oblong incubation patches, one on either side of midline (Gratto-Trevor 1992; Wiebe and Bortolotti 1993).
Incubation Period
Usually 19-21 d, occasionally up to 22 d (Safriel 1971, Jehl 2004a, Jehl 2007).
Parental Behavior
Males and females incubate equally with males usually present during the day (between 07:00 and 20:00) and females at night (20:00-07:00) (Jehl 2007), although Hitchcock and Gratto-Trevor (Hitchcock and GrattoTrevor 1997) reported no such sexual division of incubation. Incubating bird rolls eggs and camouflages nest (by bending vegetation over nest), while remaining alert. Incubation bouts 3–5 h early in incubation, and gradually increase up to 13–14 h (Ashkenazie and Safriel 1979). Daily nest attendance can vary from 80%–100% during a day, being lowest in afternoon. When one adult was experimentally removed, attendance dropped to 35% in morning and 60% in afternoon. Remaining parent almost always deserted before hatching (Erckmann 1981). If one adult is killed by a predator before eggs pip, remaining parent almost always deserts within five days (Gratto-Trevor 1991).
With both parents present, mean number of absences per day was seven (range 4–13), with mean duration of absence 21 min (2–164) in Alaska (Erckmann 1981). At Churchill, when not incubating, the sandpipers left their breeding territories and departed for mudflats 2 – 3 km distant or flew inland to feed on mudflats in a lake bordering the colony (Jehl 2006).
Hardiness Of Eggs
Embryonic resistance to chilling is maximal before incubation (Norton 1972). Tolerance to cold exposure is substantial even during incubation, but hatchability of eggs can be affected if chilling is prolonged, especially late in incubation (Erckmann 1981). When fifth eggs were experimentally introduced, they failed to hatch unless the shape of the nest cup was changed (Safriel 1980); otherwise, some eggs broke and were removed by parents.
Partial clutch loss is likely caused by mechanical damage to eggs rather than nest predation; damaged eggs in these cases are eggs of the host and not the experimentally-added eggs (Sandercock 1997b). Configuration of four pyriform eggs appears most efficient in retaining heat during incubation (Norton 1972).
Hatching
Preliminary Events And Vocalizations
Shortly before hatching, once eggs begin to pip and chicks can be heard peeping and tapping on the shell with egg teeth, parental behavior changes. Incubating bird usually sticks closer to nest, displaying and vocalizing more readily when disturbed (CLG-T). In Alaska, nest predation increases as the breeding season progresses (Sandercock 1998b).
Shell Breaking And Emergence
Chicks can hatch at any time of day. Duration of hatching of egg from first star pip to damp chick can be more than three days or less than two, but usually two to three days. Once egg has “hole-pipped,” the chick hatches in < 12 h. All eggs of a clutch usually hatch within one day, occasionally two (Ashkenazie and Safriel 1979, CLG-T). Parents rarely desert a pipped egg even if all others have hatched, and often remain incubating even unpipped eggs for several days after remainder have hatched (CLG-T).
Parental Assistance And Disposal Of Eggshells
Parents not known to assist hatching of chicks. To prevent egg-capping, eggshells sequentially removed from nest cup as young emerge from the eggs; shells flown at least 20 m from the nest by parents (Sandercock 1996). If shell crushed when egg is pipping, it is not removed by parent and chick dies. Following hatch, both parents stay with brood for first 1-2 d. Female leaves before day 6 and males take the major role in rearing the chicks from hatching to fledging, remaining with chicks until about day 14 (Jehl 2006,Jehl 2007).
Young Birds
Condition At Hatching
Average mass (± SE) of day-of-hatch chicks in Manitoba = 4.4 ± 0.04 g (n = 114); wing length 12.9 ± 0.13 mm; bill length (culmen) 7.8 ± 0.05 mm; tarsus 19.6 ± 0.08 mm; no remiges visible. Wing about 13% of adult, bill 38%, weight 16%, and tarsus 89% (CLG-T).
Covered in down at birth, with irregular dark brown markings on head and “hour-glass” shaped figure on back (Jehl 1968b). Natal down white on forehead, sides of head, and all underparts; faintly washed with pale buff on upper breast and cheeks. Median stripe on forehead partway to bill, broad loral stripe, malar stripe, and spot in center of crown are all black. Back, rump, wings and thighs a mix of hazel and black, finely dotted with white terminal tufts (Bent 1927). Legs and bill black, bill bluntly tipped, eyes open at hatching. Egg tooth lost in first day after hatching (CLG-T).
Within a few hours of hatching, chicks stumble from nest cup and peck for insect food (Ashkenazie and Safriel 1979).
Growth And Development
Mass increases rapidly until about 11th day after hatch, then more slowly (Safriel 1975, CLG-T). Bill also grows fastest in first 11 D, then more gradually. Wing length increases slowly until about day 7 (when primaries begin to appear), then rapidly to day 20, and again more slowly. Tarsus increases slowly to full adult size (CLG-T).
Primary pin feathers visible about third day after hatching. Primaries, secondaries, tertials, scapulars, wing coverts, and upper back all in pin stage by day 5 or 6. Head and breast in pin stage by day 8, and flight feathers begin to emerge. By day 10, tail and belly in pin stage, and by day 15, primaries sufficiently emerged to allow flight. By this time down present only on back of neck and head, under throat and by bill. Black rump stripe and white throat now evident. Primaries about 90% emerged by day 22, little down present, and Juvenile plumage nearly complete (CLG-T).
At day one, young birds preen, exercise wings, and crouch when warned by parents (Ashkenazi and Safriel 1979). As chicks age, they are more likely to run than crouch, and swim readily if pursued. Chicks vocalize when chilled, especially when very young. No evidence of intersibling conflict (CLG-T).
Parental Care
Brooding
Young feed themselves within hours of hatching, but are often brooded by parent for first week, and guarded constantly for first two weeks. For first 4–5 d after hatching, chicks are brooded every 3–7 min for 3–7 min (Figure 4), and males provide new scrape each night for brooding. Amount of brooding decreases substantially in second week (Safriel 1971, Ashkenazie and Safriel 1979Ashkenazie and Safriel 1979, Ashkenazi and Safriel 1979). In Alaska, adults spent 70% of time brooding and guarding chicks, and only 30% foraging, when young < 7 days; when chicks 10 days old, 70% of parental time spent foraging and only 30% in alertness, distraction displays, and brooding (Erckmann 1981).
Females normally desert brood 0–11 d after hatching, earlier if nest hatches late in season and later if nest is early. In Manitoba, 86–97% (average 91%) of females desert mates each year after hatching; in remaining 3–14% (average 9%), male deserts female. In Alaska, all females apparently desert; one was observed being chased away by mate. Females may have an energy deficit after laying, and desertion of brood may increase female survival. Males desert brood shortly before or after fledging, on average 8 d after females (Safriel 1971, Ashkenazie and Safriel 1979Ashkenazie and Safriel 1979, Ashkenazi and Safriel 1979, Gratto-Trevor 1991).
Average survival of young higher in normal 4-chick broods compared to experimental 5-chick broods if nest cup not enlarged; no significant difference in survival when nest cup is enlarged for the extra egg (Sandercock 1997b). Nonetheless, single attending parent is apparently less able to care for more than four young (Safriel 1975, Sandercock 1997b).
Juvenile females start flocking in first week of July on the natal grounds and males by 10-14 July (Jehl 2004a).
Feeding
Young not fed by parents. Chicks forage by pecking at small adult dipteran insects when very young, and mostly insect larvae when older (Holmes and Pitelka 1968).
Nest Sanitation
Chicks desert nest within a day of last chick hatching (Ashkenazie and Safriel 1979, CLG-T).
Parental Carrying Of Young
Never observed.
Cooperative Breeding
Never observed.
Brood Parasitism by Other Species
Not reported. One nest with 6 eggs, including 2 Red-necked Phalarope eggs, was incubated by Semipalmated Sandpipers (one a yearling); not known which eggs were laid first (Gratto et al. 1983).
Fledgling Stage
Departure From The Nest
Parents lead chicks from nest within hours of hatching. In Manitoba, if nesting on a small island, birds may remain until near fledging. If on a large island or mainland, parents may move chicks up to 1 km to larger water bodies, particularly coast (CLG-T). In Alaska, brood usually moved after female leaves, and second home range established up to 3 km away (Ashkenazie and Safriel 1979).
Growth
Young fly slightly 14–15 d after hatching; capable of sustained flight at 16–19 d (Safriel 1975, Gratto-Trevor 1991). At fledging, young have 80% of adult wing length, 90% bill length, 100% tarsus length, and 90% mass (CLG-T).
Association With Parents Or Other Young
See Parental care. Brood usually remains together until deserted by parents. Chicks may be adopted by other parents if broods mix (CLG-T).
Ability To Get Around, Feed, And Care For Self
Young deserted by both parents as early as 10 days of age have been known to fledge successfully (CLG-T).
Immature Stage
Juveniles migrate southward several weeks after most adults. By mid-August through September, flocks of juveniles begin arriving at staging areas along the east coast of Canada and the United States (Harrington and Morrison 1979; Hicklin 1987; Mawhinney et al. 1993). Following a short stopover and deposition of lipid energy reserves, they fly non-stop to Caribbean countries and Suriname, French and British Guiana (Rodrigues 2000; Spaans 2003).
In Suriname, once they have regained weight soon after their arrival, they undergo Prebasic molt of body feathers and some flight feathers (see Appearance: molts and plumages). Approximately two-thirds of all juveniles do not migrate northward to breed as yearlings, but spend the boreal summer on wintering grounds (Gratto and Morrison 1981, Spaans 2003Rodrigues 2000). In a northern breeding area near Barrow, AK, individuals apparently do not breed as yearlings (Safriel 1971, Safriel inMyers 1981b), but in an area 1400 km farther south, near Churchill, Man., 1%–10% of all breeders each year are yearlings (Gratto and Cooke 1987).

Arctic National Wildlife Refuge, Alaska. Photo by D. Plummer.; photographer Doug Plummer

Semipalmated Sandpiper. Unalakleet, Alaska. 8 Jun 1954 (nest and eggs); photographer WFVZ

Arctic National Wildlife Refuge, Alaska. Photo by D. Plummer.; photographer Doug Plummer

Collected at Port Manvers, Labrador, June 1912; from collections of the Field Museum of Natural History, Chicago, IL. (scale shown in cm.) Photo by P. Lowther.; photographer Peter Lowther

collected at Port Manvers, Labrador, June 1912; from collections of the Field Museum of Natural History, Chicago, IL. Photo by P. Lowther.; photographer Peter Lowther

Semipalmated Sandpiper. Unalakleet, Alaska. 8 Jun 1954 (nest and eggs); photographer WFVZ

Photo by D. Plummer.; photographer Doug Plummer

















