

Species names in all available languages
| Language | Common name |
|---|---|
| Asturian | Grña canadiana |
| Basque | Kurrilo kanadarra |
| Bulgarian | Канадски жерав |
| Catalan | grua del Canadà |
| Chinese | 沙丘鶴 |
| Chinese (SIM) | 沙丘鹤 |
| Croatian | kanadski ždral |
| Czech | jeřáb kanadský |
| Danish | Prærietrane |
| Dutch | Canadese Kraanvogel |
| English | Sandhill Crane |
| English (United States) | Sandhill Crane |
| Faroese | Kanadatrani |
| Finnish | hietakurki |
| French | Grue du Canada |
| French (France) | Grue du Canada |
| Galician | Grou americano |
| German | Kanadakranich |
| Greek | Αμερικανικός Σταχτογερανός |
| Hebrew | עגור קנדי |
| Hungarian | Kanadai daru |
| Icelandic | Mótrana |
| Japanese | カナダヅル |
| Korean | 캐나다두루미 |
| Lithuanian | Kanadinė gervė |
| Mongolian | Элсэн тогоруу |
| Norwegian | kanadatrane |
| Polish | żuraw kanadyjski |
| Portuguese (Portugal) | Grou-americano |
| Romanian | Cocor canadian |
| Russian | Канадский журавль |
| Serbian | Kanadski ždral |
| Slovak | žeriav plavokrký |
| Slovenian | Kanadski žerjav |
| Spanish | Grulla Canadiense |
| Spanish (Cuba) | Grulla Gris |
| Spanish (Mexico) | Grulla Gris |
| Spanish (Spain) | Grulla canadiense |
| Swedish | prärietrana |
| Turkish | Kanada Turnası |
| Ukrainian | Журавель канадський |
Antigone canadensis (Linnaeus, 1758)
Definitions
- ANTIGONE
- antigone
- canadensis
The Key to Scientific Names
Legend Overview
Sandhill Crane Antigone canadensis Scientific name definitions
Version: 1.0 — Published March 4, 2020
Text last updated August 17, 2014
Distribution
Introduction
Sandhill Cranes have traditionally been delineated as populations distinguished by breeding areas. For clarity, non-migratory populations are described first, followed by breeding and then wintering areas of migratory populations. Populations and descriptions therein are organized from east to west regardless of subspecies because subspecies can overlap in distribution on breeding areas, stopover sites, wintering areas, or all three. Six subspecies are recognized (see Systematics): Cuban (A. c. nesiotes), Florida (A. c. pratensis), Mississippi (A. c. pulla), Greater (A. c. tabida), Lesser (A. c canadensis), and Canadian (A. c. rowani). To minimize confusion in discussing populations vs. subspecies, Latin nomenclature is used for subspecies names throughout this account.
It is reasonable, although speculative, to hypothesize that breeding ranges of both migratory and non-migratory Sandhill Cranes were once mostly contiguous (Johnson et al. 2005a). Presumably, continent-wide reductions in numbers and distribution of cranes over the last two centuries caused populations to become fragmented into mostly discrete ranges. With numbers increasing, cranes are now expanding into areas where breeding was previously unrecorded, thus blurring separation among longstanding populations. Recent technological advances in satellite telemetry also indicate that at least a few individuals move between adjacent populations (Krapu et al. 2011; see Priorities for Future Research).
eBird data provide detailed looks at the range of this species throughout the year: eBird Year-round Range and Point Map for Sandhill Crane.
Breeding Range: Non-Migratory Populations
The three non-migratory populations are separate and contain only a single subspecies each: Cuban Population, Florida Population, and Mississippi Population (Jones et al. 2005b, Jones et al. 2005d).
Cuban Population. The Cuban Population consists of the subspecies A. c. nesiotes, the only crane species that occurs in the Caribbean; no cranes occupy Central or South America. This non-migratory population is isolated on mainland Cuba and nearby islands. Occupied areas appear locally isolated in six provinces, Isle de Juventud, Camagüey, Ciego de Avila, Sanci Spiritus, Matanzas, and Pinar del Rio (Aguilera 2002, Galvez Aguilera and Chavez-Ramrez 2010). See eBird data.
Florida Population. The Florida Population consists of the subspecies A. c. pratensis. This non-migratory population ranges from peninsular Florida as far south as Everglades National Park, and as far north as se. Georgia (Walkinshaw 1973, Stys 1997). Large concentrations of birds of the Florida Population live on the Kissimmee and Desota prairies in Florida, and the Okefenokee Swamp in Georgia (Bennett 1989b, Nesbitt and Williams 1990). See eBird data.
Mississippi Population. The Mississippi Population consists of the endangered subspecies A. c. pulla (Endangered Species Act 1973). This non-migratory population only occurs in Jackson Co., MS, between the Pascagoula River and the Harrison-Jackson county line (Aldrich 1972, Valentine and Noble 1970). Birds of the Mississippi Population primarily live on or near the Mississippi Sandhill Crane National Wildlife Refuge.
Breeding Range: Migratory Populations
Migratory Sandhill Cranes are widely distributed throughout North America, extending from the Great Lakes region to w. Alaska, and beyond North America to nw. Siberia. The species' breeding distribution extends from Baffin I., south from Mackenzie and British Columbia to s. British Columbia, and from Michigan to the Dakotas, west to Colorado, Utah, and ne. California. Recent colonization has extended the range of the species from New Jersey north to Maine. The six migratory populations are defined following breeding area affiliations. These populations are the Eastern Flyway Population, Mid-Continent Population, Rocky Mountain Population, Lower Colorado River Valley Population, Central Valley Population, and Pacific Flyway Population (Jones et al. 2005b, Jones et al. 2005d). Despite the proximity of the Mid-Continent, Central Valley, and Rocky Mountain populations, there is no evidence of interbreeding or movement among them (Drewien et al. 2000, Krapu et al. 2011).
Eastern Flyway Population. The Eastern Flyway Population consists of the subspecies A. c. tabida. This population has been increasing in numbers and distribution at least since the early 1980's (Kruse et al. 2011) with recent reoccurrence or first nesting reported in Maine, Vermont, Massachusetts, New Jersey, New York, Pennsylvania, Ohio, Illinois, Iowa, Nebraska, South Dakota, and North Dakota (Greenberg 1980, Poggensee 1992, Castrale and Bergens 2001, Melvin 2002, Ohio Department of Natural Resources 2006, Melvin 2008, Sauer et al. 2012).
Though referred to as the Eastern Flyway Population, the annual migratory movements of these individuals include periods within the Mississippi and Atlantic North American flyways. There is also likely overlap in the breeding distribution of the Eastern Flyway Population and the Mid-Continent Population (described below) in Quebec, Ontario, and Minnesota (Krapu et al. 2011), making the delineation of these populations unclear (Ad Hoc Eastern Population Sandhill Crane Committee, 2010). This is particularly true since genetic studies have confirmed occasional gene flow between these two populations (Jones et al. 2005b, Jones et al. 2005d).
The largest proportion of the population nests in the Great Lakes states region of the USA and in se. Canada, primarily Ontario, Michigan, Indiana, and Wisconsin (Ad Hoc Eastern Population Sandhill Crane Committee 2010). Though nesting records in s.-central Ontario were few in the 1980s (Tebbel 1981, Tebbel and Ankney 1982), more recent surveys suggest a growing population throughout available habitat (Bird Studies Canada 2006; http://www.birdsontario.org/atlas/maps.jsp). The Eastern Flyway Population in Wisconsin is largely located in the s.-central part of the state (Su et al. 2004; https://www.uwgb.edu/birds/wbba/species/maps/SACR.htm).
Eastern Flyway Population birds also breed in n. Illinois, n. Iowa, and se. Minnesota (Meine and Archibald 1996a). Minnesota's breeding Sandhill Cranes were clearly delineated in the 1970's (Henderson 1978), but are now loosely split between the Eastern Flyway Population in e.-central Minnesota and the Mid-Continent Population in nw. Minnesota (Tacha and Tacha 1985, Drewien and Lewis 1987). In 1992, when the last Birds of North America species account for Sandhill Crane was published, 1,200 nesting pairs of cranes were observed in these two areas. Today, the Minnesota Department of Natural Resources (2012) reports up to 5000 birds of the Eastern Flyway Population and a minimum of 7000 birds of the Mid-Continent Population; see also http://www.mnbba.org/blockmap/cresults.php?species=Sandhill%20Crane.
Mid-Continent Population. The Mid-Continent Population consists of all three migratory subspecies (A. c. tabida, A. c. canadensis, A. c rowani). This population includes birds breeding in northern temperate, subarctic, and arctic regions of North America, and despite the population name, birds breeding in eastern Asia. Thus, breeding occurs in Canada, the United States, and Russia (Johnson and Stewart 1973; Krapu et al. 2011). Within North America, the population breeds from n. Minnesota, across central Canada (from Hudson Bay to Canadian Rockies and n to Nunavut and Northwest Territories), to Alaska, and e. Siberia (Jones et al. 2005a). This population's annual cycle spans the Mississippi, Central, and Pacific North American flyways. Within the Mid-Continent Population, the Yukon Delta in Alaska has the highest nesting density of breeding pairs with densities up to 0.54 (1975) and 0.78 nests/km2 (Boise 1976, Conant et al. 1985, Melvin et al. 1990, Krapu et al. 2011). See eBird data for A. c. tabida/A. c rowani. A small population breeds in ne Russia, including the Anadyr' basin and Kamchatka Peninsula, and from Yakutia to Kolyma and Yana.
A. c. canadensis, presumably from the Mid-Continent Population, have occasionally been identified outside their normal range. At least one case is known of a juvenile, possibly through premature separation from its group or disoriented dispersal, having been observed in Florida. This individual was identified from afar by its body size, bill size, and the color of its primary feathers, but was never measured in hand (Nesbitt 1992b). Additional Sandhill Cranes thought to be A. c. canadensis from the Mid-Continent Population outside of their normal range have occurred in Rhode Island (Brewster 1890), South Carolina (Wayne 1894), and Mississippi (Nestbitt, 1992a). See eBird data for A. c. canadensis.
The Mid-Continent Population is managed in two subpopulations, the Gulf Coast subpopulation and the Western subpopulation; the two subpopulations are roughly divided by migration route and wintering area with cranes wintering along the Gulf Coast identified as the Gulf Coast subpopulation, and cranes wintering inland and west of the Gulf Coast identified as the Western subpopulation (Tacha and Vohs 1984, Tacha et al. 1994). There is known genetic exchange between these two subpopulations (Krapu et al. 2011), and cranes breeding within the breeding range of the Gulf Coast subpopulation spend 28% of fall and 42% of winter within the fall and wintering ranges of the Western subpopulation. This indicates limited biological support for the designation of the two subpopulations (Krapu et al. 2011). Based on data from 153 satellite-tagged cranes, the Mid-Continent Population may be more meaningfully defined by a total of four sub-populations according to breeding area affiliation, migratory flyway, and wintering grounds (Krapu et al. 2011).
Krapu et al. (Krapu et al. 2011) suggest the following four subpopulations: East-central Canada-Minnesota, West-central Canada-Alaska, Northern Canada-Nunavut, and Western Alaska-Siberia. Space use by the four proposed subpopulations indicates the probability of mixing on fall staging areas is low, except for the n. Canada-Nunavut and w. Canada-Alaska subpopulations (Krapu et al. 2011), though mixing on wintering grounds is decidedly likely.
East-central Canada-Minnesota. This subpopulation is proposed to include Sandhill Cranes breeding in w. Quebec, Hudson Bay Lowlands near James Bay, nw. Minnesota, n. Ontario, and the Interlake region of central Manitoba. The majority of birds breeding in this area are thought to be A. c. rowani, with some A. c. tabida (Krapu et al. 2011).
West-central Canada-Alaska. This subpopulation is proposed to include Sandhill Cranes breeding in central Saskatchewan, central and n. Alberta, ne. British Columbia, the Great Slave Plains in the Northwest Territories, and e.-central Alaska. The majority of these birds are thought to be A. c. rowani, fewer of the other two migratory subspecies.
Northern Canada-Nunavut. This subpopulation is proposed to include cranes breeding in Nunavut along the edge of the Arctic Ocean through the Boothia Peninsula, including islands of the Canadian Archipelago (i.e., Richards, Banks, and Victoria islands), westward from the northwest side of the Hudson Bay to the Yukon Territory. The majority of these birds are believed to be A. c. canadensis, with some A. c. rowani (Krapu et al. 2011).
Western Alaska-Siberia. This subpopulation is proposed to include cranes nesting from interior to coastal Alaska and westward across the Bering Sea into Russia. Cranes of the w. Alaska-Siberia subpopulation breed primarily from the Yukon-Kuskokwim Delta northward to the Seward Peninsula, as well as in ne. Russia, including the Anadyr' basin and Kamchatka Peninsula, and from Yakutia to Kolyma and Yana (Vladimirtseva et al. 2009).
The western boundary of nesting A. c. canadensis lies near the Republican Resource Reserve, Kytalyk, in Allaikh ulus of Yakutia along the Indigirka River (Vladimirtseva et al. 2009). Within these boundaries, Sandhill Cranes nest in ne. Siberia (Krapu et al. 2011), the Chyukotski Peninsula, Wrangel I., and the coast of the Arctic Ocean west to the Yana River drainage (Labutin and Degryavev 1988). Large numbers of cranes nest along one of Chukotka's largest lakes, Lake Krasnoye, which borders the Ust-Tanyurersky Regional Wildlife Refuge.
The numbers and distribution of Sandhill Cranes nesting in ne. Siberia have been increasing over the last several decades, and the range of Sandhill Cranes now overlaps that of the endangered Siberian Crane (Leucogeranus leucogeranus; Watanabe 2006). These birds are thought to be primarily A. c. canadensis, with only some A. c. rowani (Krapu et al. 2011).
Rocky Mountain Population. The Rocky Mountain Population consists exclusively of the subspecies A. c. tabida (although A. c. tabida also occur in other populations) breeding throughout central and w. Montana, south through central and e. Idaho, w. and central Wyoming, n. and central Utah, and nw. Colorado (Drewien and Bizeau 1974, Drewien et al. 2000). There is also evidence of some birds of the Rocky Mountain Population breeding in sw. Alberta (Drewien et al. 2000). This population's annual cycle spans the Central and Pacific North American flyways, although these individuals are not part of the Pacific Flyway Population.
Grays Lake NWR in se. Idaho supported the highest concentration of breeding pairs in the Rocky Mountain Population during the 1970's-1980's (Drewien and Bizeau 1974, Drewien 1973). Local abundance increased by 34% from 1970-1971 ( = 549 cranes; Drewien 1973) to 1998-2000 (735 cranes), then more recently declined to a local population of 588 cranes in 2006 (Austin et al. 2007). Though only 200 km separate the Rocky Mountain Population and Central Valley Population at their closest point, there is no known movement or pairing of A. c. tabida between these adjacent populations (Drewien et al. 2000). Similarly, movement between the Rocky Mountain Population and Lower Colorado River Valley Population is unknown, although the two groups also occur as close as 200 km of one another (Drewien et al. 2000).
Lower Colorado River Valley Population. This population is comprised of the subspecies A. c. tabida breeding primarily in ne. Nevada (Elk Co.), but also in limited locations in Idaho, Oregon, and Utah (Kruse et al. 2012). This is the smallest of all migratory Sandhill Crane populations, in both numbers and distribution. The highest breeding density occurs in the Ruby Valley, the Independence Valley, and the Upper North Fork Drainage of ne. Nevada (Laca et al. 2008).
Central Valley Population. The Central Valley Population consists of at least the subspecies A. c. tabida breeding in Nevada, California, Oregon, Washington, and British Columbia (Littlefield et al. 1994, Pogson and Lindstedt 1991, Pacific Flyway Study Committee 1997). Some members of the subspecies A. c. rowani nesting along the coasts of British Columbia and s. Alaska (Campbell et al. 1990) may also be part of the Central Valley Population, although they might be better included in the Pacific Flyway Population (Pogson and Lindstedt 1991, Ivey et al. 2005).
Members of Central Valley Population live their entire lifecycle within the Pacific Flyway, but are thought to overlap with the Pacific Flyway Population only on the wintering grounds. The Central Valley Population is divided into two unconnected subpopulations based on breeding affiliation. The northern subpopulation breeding in British Columbia, including the areas of Chilcotin Plateau, Cariboo Basin, Fraser Lowlands, n. Okanogan Valley, East Kootenay Trench, and near Vancouver is likely a mixture of A. c. tabida and A. c. rowani (Littlefield and Thompson 1979, Cooper 1996, Ivey et al 2005). The southern subpopulation breeding in s.-central Washington, e. and central Oregon, ne. California, and nw. Nevada (Littlefield and Thompson 1979, Littlefield et al. 1994, Ivey and Herziger 2000) is likely A. c. tabida.
The greatest numbers of the Central Valley Population breed in e. and central Oregon on the Malheur National Wildlife Refuge, Silvies River floodplain, and Sycan Marsh. Other major nesting concentrations occur in Klamath and Lake counties of Oregon (Littlefield et al. 1994). In California, large numbers breed in Modoc and Lassen counties (Littlefield 1989), with fewer birds in Siskiyou, Plumas, Shasta and Sierra counties (Littlefield et al. 1994, Ivey and Herziger 2000).
Non-breeding cranes congregate in common breeding areas in Oregon (Diamond Valley, Klamath Marsh National Wildlife Refuge, Warner Basin, Sycan Marsh, and the Chewaucan marshes) and California (Big Valley, Surprise Valley, and within Modoc Co.; Stern et al. 1987).
Pacific Flyway Population. The Pacific Flyway Population is comprised of at least the subspecies A. c. canadensis, and possibly A. c. rowani nesting in coastal British Columbia and Alaska. Unlike the populations described above, which occur predominantly in one North American flyway but overlap with others, the Pacific Flyway Population stays entirely within the Pacific North American flyway. Nevertheless, there may be some genetic exchange between the Pacific Flyway Population and Mid-Continent Population because their breeding areas potentially overlap in Alaska and Canada (Subcommittee on the Pacific Flyway Population of Lesser Sandhill Cranes of the Pacific Flyway Study Committee 1983).
A. c. canadensis of the Pacific Flyway Population breed in Alaska, within the upper Cook Inlet, from the Kenai Peninsula to the Alaskan Peninsula, and around Bristol Bay (Pogson 1987, Jones et al. 2005b). Cranes breeding in the Yukon-Kuskokwim Delta may be part of this population, or may be part of the Central Valley Population (Conant et al. 1981). A. c. rowani breeding along the coastal region of British Columbia, north of Vancouver I. (Cooper 2006), far into the Alexander Archipelago of se. Alaska (Willet 1921, Ivey et al. 2005), and reported on Kupreanof, Kuiu, and Sergief islands in se. Alaska (Gabrielson and Lincoln 1959), and in the Kootenay Trench region of extreme se. British Columbia could belong to either the Pacific Flyway Population or the Central Valley Population.
Nonbreeding Range
Eastern Flyway Population. Until the 1980's most of the Eastern Flyway Population of Sandhill Cranes wintered in peninsular Florida and se. Georgia (Walkinshaw 1973, Lewis et al. 1977, Crete and Toepfer 1978). The number of cranes wintering in Florida has recently declined, while numbers wintering in Georgia and other Southeastern states have increased, such that there are now a few thousand wintering in Louisiana (J. Linscombe, unpubl. data). Recent wintering has shifted north with several hundred to a few thousand Sandhill Cranes now wintering in Indiana (Castrale and Bergens 2001), e. Tennessee (i.e. at Hiwassee Wildlife Refuge, by the late 1990s wintering cranes exceeded 10,000 and by 2009 numbered around 14,000; Aborn et al. 2008, Kruse et al. 2012), and Kentucky (e.g., Barren River Lake State Resort Park; Kruse et al. 2012). Some birds even stay along s. Ontario on Lake Erie (Ad Hoc Eastern Population Sandhill Crane Committee 2010). These shifts may be in response to changes in agricultural practices or regional climate.
Mid-Continent Population. The Mid-Continent Population winters in the plains and along the Gulf Coast of Texas and Louisiana (Tacha et al. 1984, Drewien et al. 1996). Satellite transmitters indicate 82% of tagged cranes of the Mid-Continent Population use wintering grounds in Texas, with the remaining birds wintering in Oklahoma, Kansas, New Mexico, Arizona, and the Mexican states of Chihuahua, Coahuila, and Tamaulipas (Krapu et al. 2011). Small numbers of cranes also winter in central Mexico, in the states of Zacatecas and San Luis Potosi (Llanes 2012, López-Saut et al. 2011).
Following the subpopulation delineation suggested by Krapu et al. (2011), the e.-central Canada-Minnesota subpopulation concentrates along the upper Texas Gulf Coast, and Tamaulipas. Individuals from the e.-central Canada-Minnesota subpopulation have used these same wintering grounds for many decades (Tacha and Tacha 1985, Drewien and Lewis 1987, Krapu et al. 2011). The w.-central Canada-Alaska subpopulation winters primarily along the lower Texas Gulf Coast, with minimal overlap (Krapu et al. 2011).
The n. Canada-Nunavut subpopulation winters in w. Texas. The w. Alaska-Siberia subpopulation has the largest spread of wintering area locations, including w. Texas, central and s. New Mexico, se. Arizona, and throughout Chihuahua and n. Coahuila. When winter weather is not severe, breeders from w. Alaska-Siberia winter at more northern latitudes, such as those found on the Queen Charlotte Is., Vancouver I. and the lower Fraser River valley (Campbell et al. 1990); this suggests overlap with the Pacific Flyway Population. The greatest overlap among all subpopulations occurs where wintering w. Alaska-Siberia and n. Canada-Nunavut subpopulations co-occur in nw. Texas.
Within New Mexico, A. c. canadensis winter primarily along the lower Pecos River Valley, with the majority at Bitter Lake National Wildlife Refuge (Walkinshaw 1949, Boeker et al. 1961, Montgomery 1997). Another major wintering area is Grulla National Wildlife Refuge in Roosevelt Co. (Boeker et al. 1961). Some cranes of the Mid-Continent Population also winter in the middle Rio Grande Valley. The majority of cranes wintering in Mexico do so in the northern highlands of Chihuahua and Coahuila (Boeker and Baer 1962, Drewien et al. 1996). The nesting range of cranes wintering in Louisiana is unknown, but likely consists of birds from the Eastern Flyway and Mid-Continent Populations (King 2008).
Rocky Mountain Population. The Rocky Mountain Population winters primarily in the middle Rio Grande Valley of w.-central New Mexico, concentrating from the Albuquerque-Los Lunas region in Bernalillo and Valencia counties south to Bosque del Apache National Wildlife Refuge in Socorro Co. (Bailey 1928, Drewien and Bizeau 1974, Drewien et al. 2000). Because >50% of the Rocky Mountain Population winters annually at Bosque del Apache, it is considered the Rocky Mountain Population's most important wintering location (Drewien and Bizeau 1974). A smaller portion of the Rocky Mountain Population winters in sw. New Mexico, within the Hatch and Uvas valleys of Sierra and Dona Ana counties, Pecos Valley, Las Vegas National Wildlife Refuge, the Deming-Columbus area in Luna Co., and the upper Gila River, in Grant Co. (Drewien and Bizeau 1974, Lewis et al. 1977, Drewien et al. 2000).
In recent winters a small but increasing number (>1,000) have wintered in w. Colorado in the Colorado and Gunnison river valleys (Drewien and Bizeau 1974, Drewien et al. 1987, 1999). Another small group of cranes winters in the Sulphur Springs Valley in Arizona. An estimated 10% of the Rocky Mountain Population winters in the Mexican states of Durango and Chihuahua; most of these birds winter in n. Chihuahua (Drewien et al. 1996). Laguna de Babicora of Chihuahua is considered the most important wintering area in Mexico. Cranes of the Rocky Mountain and Mid-Continent populations overlap on wintering grounds, but there is no known pairing of birds across populations (Krapu et al. 2011).
Lower Colorado River Valley Population. This population winters along the lower Colorado River in w. Arizona, the Gila River in sw. Arizona, and in California's Imperial Valley (Lewis et al. 1977, Drewien et al. 1987); 90% of this population is believed to winter in and around Cibola National Wildlife Refuge in sw. Arizona, Sonny Bono Salton Sea National Wildlife Refuge in s. California, and along the Gila River in sw. Arizona (Kruse et al. 2012, Drewien et al. 1976, Rawlings 1992).
Central Valley Population. The Central Valley Population shares wintering areas with the Pacific Flyway Population in the Central and Imperial valleys of California (Lewis et al. 1977, Littlefield and Thompson 1979). Much of the population is thought to use the Sacramento Valley south to the San Joaquin-Sacramento Rivers Delta (Pogson and Lindstedt 1991), concentrating near Chico, south to Delano (Littlefield and Thompson 1979). Some 70% of A. c. tabida of the Central Valley Population winter in the Sacramento Valley (n. Central Valley) from Gridley north to near Chico, with another wintering concentration on the San Joaquin-Sacramento Rivers Delta SSW of Sacramento. During unfavorable conditions some individuals retreat south to the Delta-Grizzly I. area of California (Subcommittee on the Pacific Flyway Population of Lesser Sandhill Cranes of the Pacific Flyway Study Committee 1983).
Specific wintering areas considered important for A. c. tabida of the Central Valley Population are Stone Lakes National Wildlife Refuge, and the Staten I. and Brack Tract of California (Ivey et al. 2003). The southern extreme of the winter range for A. c. tabida is Pixley National Wildlife Refuge, NW of Delano, CA. Some individuals may winter on Sauvie I. and in neighboring sw. Washington with members of the Pacific Flyway Population. A few individuals have wintered as far north as coastal British Columbia (Littlefield et al. 1994), likely of the subspecies A. c. rowani.
Pacific Flyway Population. All members of this population winter in the Central Valley of California. Merced is thought to be the most important wintering area in the Central Valley, supporting large concentrations of cranes (Subcommittee on the Pacific Flyway Population of Lesser Sandhill Cranes of the Pacific Flyway Study Committee 1983). Large concentrations also occur in San Joaquin Co., extending south into Tulare Co. (formerly Carrizo Plains, San Luis Obispo Co.). A. c. canadensis winter primarily in the San Joaquin Valley (s. Central Valley) and on the San Joaquin River Delta. About 12% of A. c. canadensis of the Pacific Flyway Population winter in the n. Sacramento Valley, primarily in two areas, a coastal area from SE of Red Bluff to SW of Chico along the Sacramento River, and an inland area from NW of Lodi south to the Carrizo Plaines SE of McKittrick, and Pixley National Wildlife Refuge NW of Delano (Littlefield and Thompson 1982, Pogson and Lindstedt 1991).
There is no known mixing in breeding areas of these two co-wintering populations (Subcommittee on the Pacific Flyway Population of Lesser Sandhill Cranes of the Pacific Flyway Study Committee 1983). Wintering A. c. rowani belonging to either the Pacific Flyway Population or Central Valley Population occur south into Butte Co. (sw. of Chico), north into Tehama Co., and if food becomes limited, may move as far south as the San Joaquin River Delta (CDL).
Historical Changes to the Distribution
Following European expansion through North America, loss of habitat due to draining wetlands for agriculture (Samson and Knopf, 1994) and uncontrolled hunting led to sharp declines in Sandhill Crane populations throughout all major breeding populations in the contiguous United States (Walkinshaw 1949; Drewien et al. 1975, Johnson 1976, Drewien and Lewis 1987; see Conservation and Management). By the 1930's, several local populations had been extirpated or reduced to very low numbers of breeding adults (Drewien and Lewis 1987, Henika 1936). Sandhill Cranes in what is now the Mid-Continent Population were extirpated from the majority of their breeding range throughout the prairie pothole region (Drewien and Bizeau 1974, Walkinshaw 1949, Johnson 1976). Damming and inundation of bottomlands exacerbated impacts by eliminating important staging areas used during fall migration (Johnson 1963, Buller and Boeker 1965).
Although populations are now increasing, the Sandhill Crane's breeding range in North America was historically more extensive. The Eastern Flyway Population, which has had the most successful recovery, is thought to have reoccupied a major part but not all of its historic range in the Midwest U.S. (Meine and Archibald 1996a), despite predicted losses of >90% of habitat in those areas (Tiner 1984). The Mississippi Population's historical distribution is unknown, but is thought to have been part of a contiguous population across the Gulf Coast Plains of Louisiana, Mississippi, and Alabama (Walkinshaw 1949, Valentine and Noble 1970, Meine and Archibald 1996a). The historic Rocky Mountain Population extended throughout the central and n. Rocky Mountains and the Great Basin, similar to today, except that the population once occupied now vacant areas in w. Colorado and nw. Arizona (Drewien and Bizeau 1974, The Subcommittee on Rocky Mountain Greater Sandhill Cranes 2007). The Mid-Continent Population once occupied areas throughout the prairie pothole region (across North and South Dakota east of the Missouri River, s. Saskatchewan, and sw. Manitoba) that are now mostly unoccupied. The lack of recolonization of these areas may have to do with high breeding site fidelity (see Breeding), and a disproportionally large, ongoing hunting harvest of the w. Canada-Alaska and e. Canada-Minnesota subpopulations (Krapu et al. 2011).
Historical wintering habitat also once included now vacant areas south to Arizona (Mearnes 1890), Baja California in Mexico, central Mexico, the Yucatan Peninsula and Lake Texaco near Mexico City (Cooke 1914, del Campo 1944, Walkinshaw 1949, Leopold 1965, Littlefield and Ryder 1968). Early accounts of Sandhill Cranes wintering in Mexico mentioned concentrations in the Northern Highlands (San Luis Potosi) south to Pueblo (El Carmen lagoon; Cooke 1914, Leopold 1965). Most were found in Chihuahua and n. Durango (Goldman and Goldman 1935, Leopold 1959, Goldman 1942). More recent counts indicate 97.4% of Sandhill Cranes wintering in Mexico occur in the Northern Highlands, with few south of Zacatecas (Drewien et al. 1996). Loss of wintering areas in e., central, and s. Mexico are thought to have been driven by changes in agricultural practices and habitat loss (Llanes 2012).
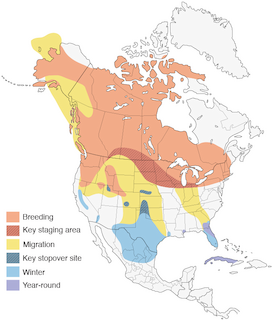
- Year-round
- Migration
- Breeding
- Non-Breeding
Northern populations are migratory, southern populations generally non-migratory, remaining near breeding sites year-round.

eBird Status and Trends

















