












Red Phalarope Phalaropus fulicarius Scientific name definitions
- LC Least Concern
- Names (61)
- Monotypic
Diane M. Tracy, Douglas Schamel, and James Dale
Version: 1.0 — Published March 4, 2020
Sign in to see your badges
Species names in all available languages
| Language | Common name |
|---|---|
| Afrikaans | Grysfraiingpoot |
| Arabic | فلروب أحمر |
| Asturian | Falaropu picugordu |
| Azerbaijani | Yastıburun üzərçə |
| Basque | Mendebal-txori mokolodia |
| Bulgarian | Плоскоклюн листоног |
| Catalan | escuraflascons becgròs |
| Chinese | 灰瓣足鷸 |
| Chinese (SIM) | 灰瓣蹼鹬 |
| Croatian | riđa liskonoga |
| Czech | lyskonoh ploskozobý |
| Danish | Thorshane |
| Dutch | Rosse Franjepoot |
| English | Red Phalarope |
| English (Kenya) | Grey Phalarope |
| English (UK) | Grey Phalarope |
| English (United States) | Red Phalarope |
| Faroese | Morlittur helsareyði |
| Finnish | isovesipääsky |
| French | Phalarope à bec large |
| French (France) | Phalarope à bec large |
| Galician | Falaropo de bico groso |
| German | Thorshühnchen |
| Greek | Κόκκινος Φαλαρόποδας |
| Hebrew | שחיינית רחבת-מקור |
| Hungarian | Laposcsőrű víztaposó |
| Icelandic | Þórshani |
| Indonesian | Kaki-rumbai merah |
| Italian | Falaropo beccolargo |
| Japanese | ハイイロヒレアシシギ |
| Korean | 붉은배지느러미발도요 |
| Latvian | Platknābja pūslītis |
| Lithuanian | Plokščiasnapis plaukikas |
| Mongolian | Улаан сэлээхэй |
| Norwegian | polarsvømmesnipe |
| Persian | فالاروپ بلوطی |
| Polish | płatkonóg płaskodzioby |
| Portuguese (Brazil) | pisa-n’água-de-bico-grosso |
| Portuguese (Portugal) | Falaropo-de-bico-grosso |
| Romanian | Notatiță cu cioc lat |
| Russian | Плосконосый плавунчик |
| Serbian | Siva liskonoga |
| Slovak | lyskonoh ploskozobý |
| Slovenian | Ploskokljuni liskonožec |
| Spanish | Falaropo Picogrueso |
| Spanish (Argentina) | Falaropo Pico Grueso |
| Spanish (Chile) | Pollito de mar rojizo |
| Spanish (Costa Rica) | Falaropo Rojo |
| Spanish (Cuba) | Zarapico rojo |
| Spanish (Ecuador) | Falaropo Rojo |
| Spanish (Mexico) | Falaropo Pico Grueso |
| Spanish (Panama) | Falaropo Rojo |
| Spanish (Paraguay) | Falaropo pico grueso |
| Spanish (Peru) | Faláropo de Pico Grueso |
| Spanish (Puerto Rico) | Falaropo Piquigrueso |
| Spanish (Spain) | Falaropo picogrueso |
| Spanish (Uruguay) | Falaropo Pico Grueso |
| Swedish | brednäbbad simsnäppa |
| Thai | นกลอยทะเลสีแดง |
| Turkish | Kızıl Denizdüdükçünü |
| Ukrainian | Плавунець плоскодзьобий |
Phalaropus fulicarius (Linnaeus, 1758)
PROTONYM:
Tringa fulicaria
Linnaeus, 1758. Systema Naturæ per Regna Tria Naturæ, Secundum Classes, Ordines, Genera, Species, cum Characteribus, Differentiis, Synonymis, Locis. Tomus I. Editio decima, reformata 1, p.148.
TYPE LOCALITY:
Hudson Bay, ex Edwards, pi. 142.
SOURCE:
Avibase, 2023
Definitions
- PHALAROPUS
- fulicaria / fulicarius
The Key to Scientific Names
Legend Overview
UPPERCASE: current genus
Uppercase first letter: generic synonym
● and ● See: generic homonyms
lowercase: species and subspecies
●: early names, variants, misspellings
‡: extinct
†: type species
Gr.: ancient Greek
L.: Latin
<: derived from
syn: synonym of
/: separates historical and modern geographic names
ex: based on
TL: type locality
OD: original diagnosis (genus) or original description (species)
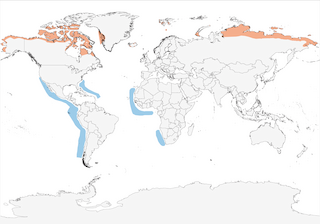
- Year-round
- Migration
- Breeding
- Non-Breeding
Distribution of the Red Phalarope















