


© Brian Small
Laysan Rail Zapornia palmeri Scientific name definitions
- EX Extinct
- Names (20)
- Monotypic
Storrs L. Olson
Version: 1.0 — Published March 4, 2020
Text last updated January 1, 1999
Text last updated January 1, 1999
Sign in to see your badges
Species names in all available languages
| Language | Common name |
|---|---|
| Catalan | rasclet de Laysan |
| Czech | chřástal laysanský |
| Dutch | Laysanral |
| English | Laysan Rail |
| English (HAW) | Moho - Laysan Rail |
| English (United States) | Laysan Rail |
| French | Marouette de Laysan |
| French (France) | Marouette de Laysan |
| German | Laysansumpfhuhn |
| Japanese | レイサンクイナ |
| Norwegian | laysanrikse |
| Polish | karliczka hawajska |
| Russian | Лайсанский погоныш |
| Serbian | Lajsanski barski petlić (izumro) |
| Slovak | chriašť laysanský |
| Spanish | Polluela de Laysan |
| Spanish (Spain) | Polluela de Laysan |
| Swedish | laysansumphöna |
| Turkish | Laysan Yelvesi |
| Ukrainian | Погонич лайсанський |
Zapornia palmeri (Frohawk, 1892)
PROTONYM:
Porzanula Palmeri
Frohawk, 1892. The Annals and Magazine of Natural History including Zoology, Botany, and Geology (6), 9, p.247.
TYPE LOCALITY:
Laysan Island, lat. 25° 46' N.; long. 171° 49' W.
SOURCE:
Avibase, 2023
Definitions
- ZAPORNIA
- zapornia
- palmeri
The Key to Scientific Names
Legend Overview
UPPERCASE: current genus
Uppercase first letter: generic synonym
● and ● See: generic homonyms
lowercase: species and subspecies
●: early names, variants, misspellings
‡: extinct
†: type species
Gr.: ancient Greek
L.: Latin
<: derived from
syn: synonym of
/: separates historical and modern geographic names
ex: based on
TL: type locality
OD: original diagnosis (genus) or original description (species)
- Year-round
- Migration
- Breeding
- Non-Breeding
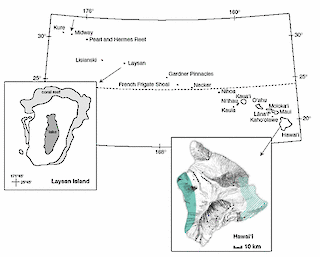
Figure 1. Former distribution of Laysan Rail endemic to Laysan Island
Former distribution of Laysan Rail endemic to Laysan I. and successfully introduced to Midway Atoll. Approximate former distribution of Hawaiian Rail on leeward side of Hawai‘i I. (shaded), so far as is known from archaeological and paleontological sites, incorporating the probable source area of the 2 mounted specimens referred to P. s. sandwichesis; and the distribution of P. s. millsi on the windward side (hatched) as reconstructed from historical accounts.















