









Great-tailed Grackle Quiscalus mexicanus Scientific name definitions
- LC Least Concern
- Names (31)
- Subspecies (8)
Kristine Johnson and Brian D. Peer
Version: 2.0 — Published January 14, 2022
Revision Notes
Revision Notes
Sign in to see your badges
Species names in all available languages
| Language | Common name |
|---|---|
| Asturian | Quiscalue mexican |
| Catalan | quíscal de Mèxic |
| Czech | vlhovec velkoocasý |
| Dutch | Langstaarttroepiaal |
| English | Great-tailed Grackle |
| English (United States) | Great-tailed Grackle |
| French | Quiscale à longue queue |
| French (France) | Quiscale à longue queue |
| Galician | Graúna mexicana |
| German | Großschwanzgrackel |
| Icelandic | Bátsóti |
| Japanese | オナガクロムクドリモドキ |
| Norwegian | storhaletrupial |
| Polish | wilgowron meksykański |
| Russian | Большехвостый гракл |
| Serbian | Dugorepi kukuruzar |
| Slovak | vlhovec člnkochvostý |
| Spanish | Zanate Mexicano |
| Spanish (Chile) | Zanate mexicano |
| Spanish (Costa Rica) | Zanate Grande |
| Spanish (Ecuador) | Clarinero Coligrande |
| Spanish (Honduras) | Zanate |
| Spanish (Mexico) | Zanate Mayor |
| Spanish (Panama) | Tordo Coligrande |
| Spanish (Peru) | Clarinero de Cola Grande |
| Spanish (Puerto Rico) | Zanate Mejicano |
| Spanish (Spain) | Zanate mexicano |
| Spanish (Venezuela) | Galandra |
| Swedish | mexikobåtstjärt |
| Turkish | Büyük Kuyruklu Kiskal |
| Ukrainian | Гракл великохвостий |
Revision Notes
Kristine Johnson revised the account, with contributions from Peter Pyle on the "Appearances" page. Peter Pyle and Tammy Zhang curated the media, and Vicens Vila-Coury generated the range map.
Quiscalus mexicanus (Gmelin, 1788)
PROTONYM:
C.[orvus] mexicanus
Gmelin, 1788. Systema Naturae. Editio decima tertia, aucta, reformata. Cura Jo. Frid. Gmelin (etc.). Tomus I [pars I], p.375.
TYPE LOCALITY:
'in nova Hispania' (cf. Peters, 1929, Proc. Biol. Soc. Washington, 42, p. 121) ; restricted to region about Veracruz, Veracruz, Mexico, by Lowery, 1938, Occas. Papers Mus. Zool. Louisiana State Univ., no. 1, p. 4.
SOURCE:
Avibase, 2023
Definitions
- QUISCALUS
- mexicanum / mexicanus
The Key to Scientific Names
Legend Overview
UPPERCASE: current genus
Uppercase first letter: generic synonym
● and ● See: generic homonyms
lowercase: species and subspecies
●: early names, variants, misspellings
‡: extinct
†: type species
Gr.: ancient Greek
L.: Latin
<: derived from
syn: synonym of
/: separates historical and modern geographic names
ex: based on
TL: type locality
OD: original diagnosis (genus) or original description (species)
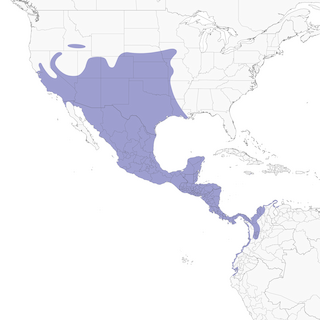
- Year-round
- Migration
- Breeding
- Non-Breeding
Distribution of the Great-tailed Grackle















