

Species names in all available languages
| Language | Common name |
|---|---|
| Catalan | bosquerola aladaurada |
| Czech | lesňáček zlatokřídlý |
| Danish | Gulvinget Sanger |
| Dutch | Geelvleugelzanger |
| English | Golden-winged Warbler |
| English (United States) | Golden-winged Warbler |
| French | Paruline à ailes dorées |
| French (France) | Paruline à ailes dorées |
| German | Goldflügel-Waldsänger |
| Greek | Χρυσόφτερη Πάρουλα |
| Haitian Creole (Haiti) | Ti Tchit zèl dore |
| Hebrew | סבכון זהוב-כנף |
| Hungarian | Aranyszárnyú hernyófaló |
| Icelandic | Gullskríkja |
| Japanese | キンバネアメリカムシクイ |
| Lithuanian | Geltonsparnis kirmlesys |
| Norwegian | gullvingeparula |
| Polish | lasówka złotoskrzydła |
| Portuguese (Portugal) | Mariquita-d'asa-amarela |
| Romanian | Omidar cu aripă aurie |
| Russian | Желтокрылая червеедка |
| Serbian | Zlatokrila cvrkutarka |
| Slovak | horárik zlatokrídly |
| Slovenian | Zlatoperuti peničar |
| Spanish | Reinita Alidorada |
| Spanish (Costa Rica) | Reinita Alidorada |
| Spanish (Cuba) | Bijirita alidorada |
| Spanish (Dominican Republic) | Cigüita Ala de Oro |
| Spanish (Ecuador) | Reinita Alidorada |
| Spanish (Honduras) | Chipe Ala Dorada |
| Spanish (Mexico) | Chipe Alas Amarillas |
| Spanish (Panama) | Reinita Alidorada |
| Spanish (Peru) | Reinita de Ala Dorada |
| Spanish (Puerto Rico) | Reinita Alidorada |
| Spanish (Spain) | Reinita alidorada |
| Spanish (Venezuela) | Reinita Alidorada |
| Swedish | guldvingad skogssångare |
| Turkish | Altın Kanatlı Ötleğen |
| Ukrainian | Червоїд золотокрилий |
Vermivora chrysoptera (Linnaeus, 1766)
Definitions
- VERMIVORA
- vermivora / vermivorum / vermivorus
- chrysoptera / chrysopterum
The Key to Scientific Names
Legend Overview
Golden-winged Warbler Vermivora chrysoptera Scientific name definitions
Version: 1.0 — Published March 4, 2020
Text last updated March 25, 2011
Demography and Populations
Measures of Breeding Activity
Age At First Breeding; Intervals Between Breeding.
Normally breeds by second year (age 10-12 mo), and every year thereafter.
Clutch
Mean values of: 5.0 ± 0.60 (SD) (n = 74) from throughout the range for nests from 1880-1930, excluding 1 nest with only 2 eggs (Western Foundation for Vertebrate Zoology). Recent surveys: 4.8 ± 0.6 (SD) (n = 12) in n.-central New York, excluding one abandoned clutch with one egg (JLC); 5.15 (n = 13) in central Michigan assuming one warbler egg lost per cowbird egg (Will 1986); 4.30 ± 0.09 (SE) (n = 102) in the Cumberland Mountains of Tennessee (Bulluch 2007); 4.95 ± 0.10 (SE) (n = 86) at Queen's University Biological Station in e. Ontario, Canada (Vallender 2006).
Second attempts at nesting usually occur if the first nest is unsuccessful. But if the first attempt is successful, production of a second clutch is unknown.
Annual Reproductive Success
Nest success rates (ns) following Mayfield are determined from daily nest survival rates (dns) raised to the 25th power including the duration of egg laying, incubation and nestling stage, or raised to the 21st power by ignoring egg laying because nests discovered during egg laying are often abandoned but rapidly restarted. Using the authors' preference for these two options: dns of 97.2% to the 25th provides ns = 48.8% (n = 102) in uplands of Cumberland Mountains, TN (Bulloch 2007); dns of 95.5% to the 25th provides ns = 32.8% (n = 86) in e. Ontario (Vallender-Fraser 2006); dns of 97.8% to the 21st provides ns = 63% (n = 44) in swamp forests in s. New York (JLC); and dns of 95.8% to the 21st provides ns = 41% (n = 32) in uplands in s. New York (JLC).
A mean annual fledging rate excluding second attempts after initial failure: 3.3/nest (n = 13) in central Michigan (Will 1986) and 2.0/nest (n = 40) in n.-central New York (JLC). Fecundity, i.e., mean annual fledging rate per female including estimated second nest attempts: 1.61 (n = 102) in Tennessee (Bulluch 2007), 1.53 (n = 86) in s. Ontario (Vallender-Fraser 2006), 1.76 (n = 32) for swamp forests in s. New York (JLC), and 1.24 for uplands in s. New York (JLC). Applying a correction factor for hybridization by assuming that young fledged by hybrid pairs reduce genetic transmission and fecundity by half, the corrected fecundity estimates are 1.67 for swamp forests in s. New York and 1.17 for uplands in s. New York.
Life Span And Survivorship
Poorly sampled. All following records based on resighting at breeding site. Two records of 7-year old males from Bird Banding Laboratory. In s. New York: one 7-year old male observed out of 28 males banded at least 6 years before end of field study; one female at least 7-year old (banded as an after second year bird) and one 6-year old female observed out of 23 females banded at least 5 years before end of field study (JLC). In n. Wisconsin, 5 of 35 (14%) males banded in 2007 were observed in 2010 (AMR).
From BBL banding records: maximum longevity 7 yr 11 mo (http://www.pwrc.usgs.gov/BBL/homepage/long5930.htm). .
Mortality And Disease
Not known.
Population Spatial Metrics
Natal Philopatry
Discovery of second year birds returning to the natal site is very low: 0 of 88 banded nestlings or fledglings (Will 1986), although nearly 3% in studies in West Virginia (Aldinger pers. comm.).
Fidelity To Breeding Sites
Males resighted at breeding sties: central Michigan, 8 of 11 (73%) (Murray and Gill 1976); central Wisconsin, 12 of 24 (50%) (Will 1986); central New York, 3 of 10 (30%) (Confer and Knapp 1979); n.-central New York, 30 of 63 (48%) and s. New York, 44 of 88 (50%) (JLC), n. Wisconsin, 45 of 97 (46%). In se. Kentucky, 5 of 8 (63%) males banded in 2004 returned to a former breeding site in 2005, with 3 of these males still present in 2008. An additional 8 of 14 (57%) males banded in 2007 returned to the same area in 2008 (PJH).
Resighting rates appear lower for females than for males in Tennessee (Bulluch 2007), s. Ontario (Vallender-Fraser 2006) and s. New York (JLC). It is not known if the lower rate is due to a difference between males and females for survival, detectability, or site fidelity, or some mix of these factors.
Fidelity To Winter Sites
Not known.
Population Status
See Table 3. The Golden-winged Warbler population has been declining for ≥40 years at 2.8% per year (Breeding Bird Survey [BBS] data for 1966-2007, Sauer et al. 2008). Because of population declines throughout much of its range, the species is listed as "threatened" by the Committee on the Status of Endangered Wildlife in Canada, is a "species of management concern" in the United States (U.S. Fish and Wildlife Service), and is on the Partners-in-Flight Continental Watch List (Rich et al. 2004). Golden-winged Warblers are state-listed as "endangered" in Indiana, Ohio, Georgia, and Massachusetts, "threatened" in Kentucky and "of management or special concern" in Tennessee, New Jersey, New York, Connecticut, and Wisconsin. Golden-winged Warblers are also listed as "extremely high priority" on the Audubon's Society's Watch List (National Audubon Society 2002).
The Golden-winged Warbler Working Group (2010) provides this overview: (A) "Golden-winged Warbler populations are declining throughout all of their range as early-successional habitats revert to forest and as upland and wetland habitats are lost to human development. These declines are resulting in extirpation of the species from areas that have supported Golden-winged Warblers for at least the last century (Georgia, South Carolina, Virginia, Massachusetts, Connecticut, Rhode Island, Vermont, New Hampshire, Indiana, Illinois, Ohio)."
(B) Paraphrased from this source for recent BBS results: The northern range in Ontario, Minnesota, Wisconsin, Michigan and Manitoba once seemed to provide a refuge for Golden-winged Warblers, but analyses of 1998-2007 suggest a rapid rate of decline in the southern portion of the northern population (4.4% annually in Ontario, 2.4% annually in Wisconsin and 2.2% annually in Michigan), as well as the long-term decline of the southern portion.
(C) "Overall, Golden-winged Warblers showed stable or increasing populations for the entire BBS period (1966-2003) in the Boreal-Hardwood Transition region and neighboring Ontario. However, analyses of the last 10 yr of BBS data (1994-2003) show an annual decline of 9.0% in the FWS Region 3 (n.-central states of Minnesota, Wisconsin, and Michigan), an 11.3% decline annually in Ontario."
(D) "The northern range once seemed to provide a refuge for Golden-winged Warblers, but analyses of recent trends suggest a very rapid rate of decline in the southern portion of the northern population as well as long-term decline of the southern portion" (see Table 3).
Population Regulation
Five commonly cited factors may contribute to regional declines of this species: (1) loss of breeding habitat; (2) Brown-headed Cowbird parasitism; (3) interference or exploitative competition with Blue-winged Warblers; (4) hybridization with Blue-winged Warblers; and (5) loss of winter habitat.
Virtually all regions where Golden-winged Warblers have been extirpated or are currently declining have lost habitat due to extensive reforestation or urban sprawl. This loss of anthropogenic disturbance habitat is an important cause of the decline (e.g., Confer and Pascoe 2003).
In n.-central New York with moderate density of Brown-headed Cowbirds (Sauer et al. 2008), nest parasitism lowered the fledging rate by 17% during a five-year survey (Confer et al. 2003). In s. New York with reduced agriculture and fewer cowbirds (Sauer et al. 2008), nest parasitism reduced fecundity by about 5% (JLC). Golden-winged Warblers expanded into New England during the 1800s (Gill 1980) when agriculture and live stock and probably cowbirds were abundant and have recently expanded into n.-central New York (Andrle and Carrolle 1988, McGowan and Corwin 2008) where cowbirds remain moderately abundant (Sauer et al. 2008). Thus, the Golden-wing population is able to overcome effects of nest parasitism if other factors are suitable.
Extirpation has commonly occurred in areas where the Blue-winged Warbler has invaded the Golden-wing range, even where suitable habitat remains and is unoccupied by either species (Confer and Pascoe 2003). In almost all areas of Blue-wing intrusion, the Golden-wing phenotype is displaced within 50 yr (Gill 1988) or less (Canterbury et al. 1993). The reciprocal displacement of the Blue-wing phenotype by the Golden-wing phenotype has never been observed. A Blue-wing competitive advantage may contribute to the Golden-wing decline (Will 1986, Confer et al. 2003), but it is hard to see this as a major factor because the Golden-winged Warbler is larger and dominates the majority of agonistic interactions (Confer and Larkin 1988), although not in all studies (Will 1986).
The influence of hybridization on the relative abundance of Golden-winged and Blue-winged Warblers is not clear. A parental cross of phenotypically pure birds would seem to be equally deleterious for both species by equally diluting their genes. Hybrids are fertile but experience negative sexual selection in some populations (Ficken and Ficken 1968, Confer and Tupper 2000, Leichty and Grier 2006, Canterbury pers. comm.), but not in s. Ontario (Vallender pers. comm.). This could occur intrasexually through male aggression toward hybrid phenotypes and/or intersexually because of female mating preferences for pure types (Leichty and Grier 2006). Back crosses may occur more frequently with Golden-winged than Blue-winged warblers: back crosses occurred with 9.1% of 231 pairings that included a phenotypically pure Golden-winged Warbler but only 5.1% of 117 pairings with a Blue-winged Warbler for the sum of observations in north central New York (Confer and Larkin 1998) and s. New York (JLC). Similarly, 9 of 9 mixed species pairs in Kentucky included a pure golden-wing phenotype while only 2 blue-wing phenotypes were found in mixed species pairs (PJH). If statistically verified, a higher frequency of hybridization by Golden-winged Warblers would help account for their replacement by Blue-winged Warblers.
The potential effect of loss of winter habitat on populations is unclear, especially because the winter range remains poorly documented. The lack of any detectable difference in apparent winter range for populations in s. New York that are declining and in s. Ontario that have recently increased provides a weak suggestion that winter habitat is not a regulatory factor.
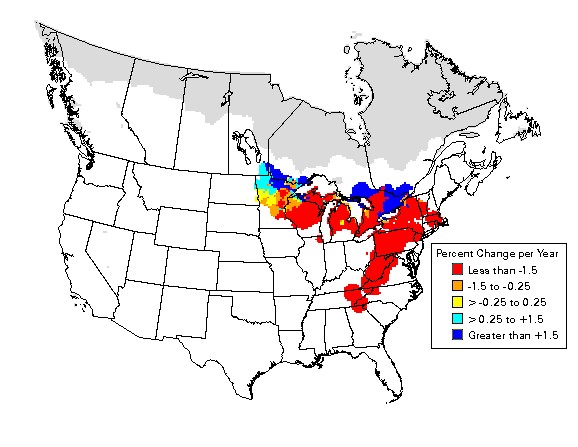
Based on data from the Breeding Bird Survey. From Sauer et al. 2008.















