












Five-striped Sparrow Amphispizopsis quinquestriata Scientific name definitions
- LC Least Concern
- Names (19)
- Subspecies (2)
Kathleen D. Groschupf
Version: 1.1 — Published August 18, 2021
Sign in to see your badges
Species names in all available languages
| Language | Common name |
|---|---|
| Catalan | sit de cinc ratlles |
| Dutch | Vijfstrepengors |
| English | Five-striped Sparrow |
| English (United States) | Five-striped Sparrow |
| French | Bruant pentaligne |
| French (France) | Bruant pentaligne |
| German | Fünfstreifenammer |
| Icelandic | Fingurtittlingur |
| Japanese | ムナフスズメモドキ |
| Norwegian | sonoraspurv |
| Polish | wąsatek białobrody |
| Russian | Пятиполосая овсянка |
| Slovak | strnádlik päťpásy |
| Spanish | Chingolo Cincorrayas |
| Spanish (Mexico) | Zacatonero Cinco Rayas |
| Spanish (Spain) | Chingolo cincorrayas |
| Swedish | fembandad sparv |
| Turkish | Beş Çizgili Serçe |
| Ukrainian | Ґоріон білогорлий |
Amphispizopsis quinquestriata (Sclater & Salvin, 1868)
PROTONYM:
Zonotrichia quinquestriata
Sclater & Salvin, 1868. Proceedings of the Zoological Society of London 1868, Part 2, p.323.
TYPE LOCALITY:
Mexico ; type probably from Bolanos, Jalisco, fide Miller, Friedmann, Griscom, and Moore, 1957, Pacific Coast Avifauna (Cooper Ornith. Soc.) , no. 33, p. 371.
SOURCE:
Avibase, 2023
Definitions
- AMPHISPIZOPSIS
- quinquestriata
The Key to Scientific Names
Legend Overview
UPPERCASE: current genus
Uppercase first letter: generic synonym
● and ● See: generic homonyms
lowercase: species and subspecies
●: early names, variants, misspellings
‡: extinct
†: type species
Gr.: ancient Greek
L.: Latin
<: derived from
syn: synonym of
/: separates historical and modern geographic names
ex: based on
TL: type locality
OD: original diagnosis (genus) or original description (species)
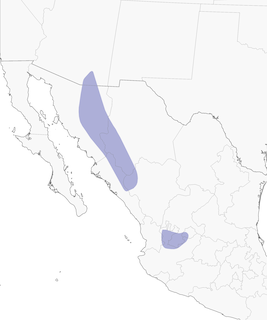
- Year-round
- Migration
- Breeding
- Non-Breeding
Distribution of the Five-striped Sparrow















