














Eared Grebe Podiceps nigricollis Scientific name definitions
- LC Least Concern
- Names (55)
- Subspecies (3)
S. A. Cullen, Joseph R. Jehl Jr., and Gary L. Nuechterlein
Version: 1.0 — Published March 4, 2020
Sign in to see your badges
Species names in all available languages
| Language | Common name |
|---|---|
| Afrikaans | Swartnekdobbertjie |
| Albanian | Kredharaku gushëzi |
| Arabic | غطاس أسود الرقبة |
| Armenian | Սևավիզ սուզակ |
| Asturian | Somorguyu de pescuezu prietu |
| Azerbaijani | Qaraboyun batağan |
| Basque | Txilinporta lepabeltza |
| Bulgarian | Черноврат гмурец |
| Catalan | cabussó collnegre |
| Chinese | 黑頸鸊鷉 |
| Chinese (SIM) | 黑颈䴙䴘 |
| Croatian | crnogrli gnjurac |
| Czech | potápka černokrká |
| Danish | Sorthalset Lappedykker |
| Dutch | Geoorde Fuut |
| English | Eared Grebe |
| English (Bangladesh) | Black-necked Grebe |
| English (India) | Eared Grebe (Black-necked Grebe) |
| English (Kenya) | Black-necked Grebe |
| English (South Africa) | Black-necked Grebe |
| English (UK) | Black-necked Grebe |
| English (United Arab Emirates) | Black-necked Grebe |
| English (United States) | Eared Grebe |
| Finnish | mustakaulauikku |
| French | Grèbe à cou noir |
| French (France) | Grèbe à cou noir |
| Galician | Mergullón de pescozo negro |
| German | Schwarzhalstaucher |
| Greek | Μαυροβουτηχτάρι |
| Hebrew | טבלן בינוני |
| Hungarian | Feketenyakú vöcsök |
| Icelandic | Stargoði |
| Italian | Svasso piccolo |
| Japanese | ハジロカイツブリ |
| Korean | 검은목논병아리 |
| Latvian | Melnkakla dūkuris |
| Lithuanian | Juodakaklis kragas |
| Mongolian | Халтар шунгуур |
| Norwegian | svarthalsdykker |
| Persian | کشیم گردن سیاه |
| Polish | zausznik |
| Portuguese (Portugal) | Cagarraz |
| Romanian | Corcodel cu gât negru |
| Russian | Черношейная поганка |
| Serbian | Crnovrati gnjurac |
| Slovak | potápka čiernokrká |
| Slovenian | Črnovrati ponirek |
| Spanish | Zampullín Cuellinegro |
| Spanish (Costa Rica) | Zambullidor Mediano |
| Spanish (Mexico) | Zambullidor Orejón |
| Spanish (Spain) | Zampullín cuellinegro |
| Swedish | svarthalsad dopping |
| Thai | นกเป็ดผีคอดำ |
| Turkish | Kara Boyunlu Batağan |
| Ukrainian | Пірникоза чорношия |
Podiceps nigricollis Brehm, 1831
PROTONYM:
Podiceps nigricollis
Brehm, 1831. Handbuch der Naturgeschichte aller Vögel Deutschlands, p.963.
TYPE LOCALITY:
Germany.
SOURCE:
Avibase, 2023
Definitions
- PODICEPS
- podiceps
- nigricollis
The Key to Scientific Names
Legend Overview
UPPERCASE: current genus
Uppercase first letter: generic synonym
● and ● See: generic homonyms
lowercase: species and subspecies
●: early names, variants, misspellings
‡: extinct
†: type species
Gr.: ancient Greek
L.: Latin
<: derived from
syn: synonym of
/: separates historical and modern geographic names
ex: based on
TL: type locality
OD: original diagnosis (genus) or original description (species)
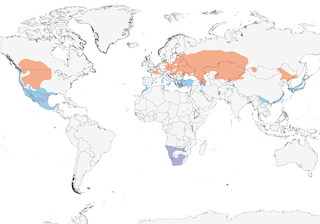
- Year-round
- Migration
- Breeding
- Non-Breeding
Distribution of the Eared Grebe















