













Cooper's Hawk Astur cooperii Scientific name definitions
- LC Least Concern
- Names (29)
- Monotypic
Robert N. Rosenfield, Kristin K. Madden, John Bielefeldt, and Odette E. Curtis
Version: 1.1 — Published October 22, 2024
Revision Notes
Revision Notes
Sign in to see your badges
Species names in all available languages
| Language | Common name |
|---|---|
| Bulgarian | Американски среден ястреб |
| Catalan | astor de Cooper |
| Croatian | crnokapi jastreb |
| Czech | jestřáb americký |
| Dutch | Coopersperwer |
| English | Cooper's Hawk |
| English (United States) | Cooper's Hawk |
| Estonian | põhja-tanukull |
| Finnish | kyyhkyhaukka |
| French | Épervier de Cooper |
| French (Canada) | Épervier de Cooper |
| German | Rundschwanzhabicht |
| Icelandic | Eikarhaukur |
| Japanese | クーパーハイタカ |
| Norwegian | trostehauk |
| Polish | krogulec czarnołbisty |
| Russian | Куперов ястреб |
| Serbian | Kuperov kobac |
| Slovak | jastrab čiapočkatý |
| Slovenian | Cooperjev kragulj |
| Spanish | Gavilán de Cooper |
| Spanish (Costa Rica) | Gavilán de Cooper |
| Spanish (Honduras) | Gavilán Cola Redondeada |
| Spanish (Mexico) | Gavilán de Cooper |
| Spanish (Panama) | Gavilán de Cooper |
| Spanish (Spain) | Gavilán de Cooper |
| Swedish | trastduvhök |
| Turkish | Cooper Atmacası |
| Ukrainian | Яструб чорноголовий |
Revision Notes
Shawn M. Billerman contributed to the Systematics section. Billy Dignam prepared the account for the 2024 Clements taxonomy update.
Astur cooperii (Bonaparte, 1828)
PROTONYM:
Falco cooperii
Bonaparte, 1828. American Ornithology or, the Natural History of the Birds of the United States not given by Wilson with figures drawn engraved and coloured from nature by Charles Lucien Bonaparte 2, p.1 pl.10 fig.1.
TYPE LOCALITY:
Near Bordentown, N. J.
SOURCE:
Avibase, 2024
Definitions
- ASTUR
- astur
- cooperi / cooperii
The Key to Scientific Names
Legend Overview
UPPERCASE: current genus
Uppercase first letter: generic synonym
● and ● See: generic homonyms
lowercase: species and subspecies
●: early names, variants, misspellings
‡: extinct
†: type species
Gr.: ancient Greek
L.: Latin
<: derived from
syn: synonym of
/: separates historical and modern geographic names
ex: based on
TL: type locality
OD: original diagnosis (genus) or original description (species)
Content Partner

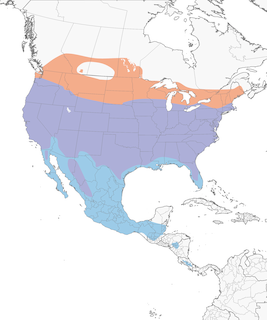
- Year-round
- Migration
- Breeding
- Non-Breeding
Distribution of the Cooper's Hawk


































