










Black-throated Green Warbler Setophaga virens Scientific name definitions
- LC Least Concern
- Names (40)
- Monotypic
Douglass H. Morse and Alan F. Poole
Version: 1.0 — Published March 4, 2020
Text last updated August 1, 2005
Text last updated August 1, 2005
Sign in to see your badges
Species names in all available languages
| Language | Common name |
|---|---|
| Catalan | bosquerola gorjanegra |
| Czech | lesňáček proužkoboký |
| Danish | Sorthalset Sanger |
| Dutch | Gele Zwartkeelzanger |
| English | Black-throated Green Warbler |
| English (United States) | Black-throated Green Warbler |
| French | Paruline à gorge noire |
| French (France) | Paruline à gorge noire |
| German | Grünmantel-Waldsänger |
| Greek | Πράσινη Πάρουλα |
| Haitian Creole (Haiti) | Ti Tchit fal nwa |
| Hebrew | סבכון צהוב-ראש |
| Hungarian | Feketetorkú lombjáró |
| Icelandic | Grænskríkja |
| Japanese | ノドグロミドリアメリカムシクイ |
| Lithuanian | Žalsvasis krūminukas |
| Norwegian | grønnparula |
| Polish | lasówka czarnogardła |
| Portuguese (Brazil) | mariquita-de-garganta-preta |
| Portuguese (Portugal) | Mariquita-verde-de-garganta-preta |
| Romanian | Omidar cu gât negru |
| Russian | Зелёная древесница |
| Serbian | Zelena crnogrla cvrkutarka |
| Slovak | horárik zelený |
| Slovenian | Črnoprsi gozdičar |
| Spanish | Reinita Dorsiverde |
| Spanish (Chile) | Reinita de dorso verde |
| Spanish (Costa Rica) | Reinita Cariamarilla |
| Spanish (Cuba) | Bijirita de garganta negra |
| Spanish (Dominican Republic) | Cigüita Pechinegro |
| Spanish (Ecuador) | Reinita Cariamarilla |
| Spanish (Honduras) | Chipe Verdoso Garganta Negra |
| Spanish (Mexico) | Chipe Dorso Verde |
| Spanish (Panama) | Reinita Negriverde |
| Spanish (Puerto Rico) | Reinita Verdosa |
| Spanish (Spain) | Reinita dorsiverde |
| Spanish (Venezuela) | Reinita Gorginegra |
| Swedish | grönryggig skogssångare |
| Turkish | Yeşil Sırtlı Ötleğen |
| Ukrainian | Пісняр-лісовик чорногорлий |
Setophaga virens (Gmelin, 1789)
PROTONYM:
Motacilla virens
Gmelin, 1789. Systema Naturae. Editio decima tertia, aucta, reformata. Cura Jo. Frid. Gmelin (etc.). Tomus I. Pars II 1 pt2, p.985.
TYPE LOCALITY:
Pennsylvania ; restricted to Philadelphia, Pennsylvania, by Amer. Ornith. Union, 1910, Checkhst North Amer. Birds, ed. 3, p. 317.
SOURCE:
Avibase, 2023
Definitions
- SETOPHAGA
- virens
The Key to Scientific Names
Legend Overview
UPPERCASE: current genus
Uppercase first letter: generic synonym
● and ● See: generic homonyms
lowercase: species and subspecies
●: early names, variants, misspellings
‡: extinct
†: type species
Gr.: ancient Greek
L.: Latin
<: derived from
syn: synonym of
/: separates historical and modern geographic names
ex: based on
TL: type locality
OD: original diagnosis (genus) or original description (species)
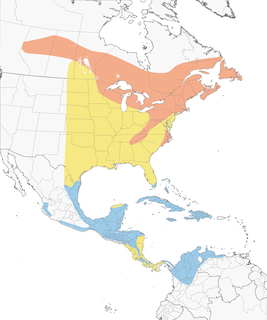
- Year-round
- Migration
- Breeding
- Non-Breeding
Distribution of the Black-throated Green Warbler















