

Species names in all available languages
| Language | Common name |
|---|---|
| Catalan | bosquerola estriada |
| Czech | lesňáček černohlavý |
| Danish | Stribet Sanger |
| Dutch | Zwartkopzanger |
| English | Blackpoll Warbler |
| English (United States) | Blackpoll Warbler |
| Finnish | viirukerttuli |
| French | Paruline rayée |
| French (France) | Paruline rayée |
| German | Streifenwaldsänger |
| Greek | Μελανοπάρουλα |
| Haitian Creole (Haiti) | Ti Tchit sèjan |
| Hebrew | סבכון פסים |
| Hungarian | Kucsmás lombjáró |
| Icelandic | Rákaskríkja |
| Japanese | ズグロアメリカムシクイ |
| Lithuanian | Dryžuotasis krūminukas |
| Norwegian | svarthetteparula |
| Polish | lasówka czarnogłowa |
| Portuguese (Brazil) | mariquita-de-perna-clara |
| Portuguese (Portugal) | Mariquita-de-perna-clara |
| Romanian | Frunzar striat |
| Russian | Пестрогрудая древесница |
| Serbian | Prugasta cvrkutarka |
| Slovak | horárik tajgový |
| Slovenian | Črnoglavi gozdičar |
| Spanish | Reinita Estriada |
| Spanish (Argentina) | Arañero Estriado |
| Spanish (Chile) | Monjita americana |
| Spanish (Costa Rica) | Reinita Rayada |
| Spanish (Cuba) | Bijirita de cabeza negra |
| Spanish (Dominican Republic) | Cigüita Cabeza Negra |
| Spanish (Ecuador) | Reinita Estriada |
| Spanish (Honduras) | Chipe Copa Negra |
| Spanish (Mexico) | Chipe Cabeza Negra |
| Spanish (Panama) | Reinita Estriada |
| Spanish (Peru) | Reinita Estriada |
| Spanish (Puerto Rico) | Reinita Rayada |
| Spanish (Spain) | Reinita estriada |
| Spanish (Uruguay) | Arañero Estriado |
| Spanish (Venezuela) | Reinita Rayada |
| Swedish | vitkindad skogssångare |
| Turkish | Kara Kırçıllı Ötleğen |
| Ukrainian | Пісняр-лісовик білощокий |
Setophaga striata (Forster, 1772)
Definitions
- SETOPHAGA
- striata
The Key to Scientific Names
Legend Overview
Blackpoll Warbler Setophaga striata Scientific name definitions
Version: 1.0 — Published March 4, 2020
Text last updated June 4, 2013
Distribution
Introduction
Breeding Range
Boreal spruce and fir forests of Alaska, Canada, and the ne. U.S. (Figure 1). Northern portion of range is a broad belt extending from Alaska (south of the Brooks Range to the base of the Alaska Peninsula; Armstrong 1980), the mouth of Mackenzie River (Yukon; Sinclair et al. 2003), and n. British Columbia (Campbell et al. 2001) east through w. and s. Mackenzie, s. Keewatin, and northern portions of Alberta (south to Banff in Canadian Rockies; Fed. Alberta Naturalists 2007), Saskatchewan and Manitoba (south to roughly 54°N), and Ontario (south to south end of James Bay; http://www.birdsontario.org/atlas/maps.jsp) to central Quebec (south of Ungava Peninsula), central Labrador, Newfoundland, the Maritime Provinces, and the ne. U.S. (Godfrey 1966, Dunn and Garrett 1997).
Locally distributed in most of its southeastern range, where limited to mountainous areas of e. New York (Adirondacks and Catskills; McGowan and Corwin 2008), ne. Massachusetts (Mt. Greylock; Veit and Petersen 1993), Vermont (Green Mtns.; Laughlin and Kibbe 1985), and New Hampshire (White Mtns.; Foss 1994a), n. and se. Maine, se. Quebec (Laurentides Provincial Park to Gaspe Peninsula; Cyr and Larivee 1995a), n. and sw. New Brunswick, and sw. and n. Nova Scotia (including Cape Breton I.) (Erskine 1992a; http://www.mba-aom.ca/jsp/map.jsp?lang=en ). Breeding areas in New York and s. Nova Scotia are fairly disjunct from the rest of the range (http://www.dec.ny.gov/cfmx/extapps/bba/bbaMaps.cfm?bndcode=BLBW&order=2&year=2000&comp=0 ).
Local breeding documented in the late 1980s and early 1990s in areas south of the main range: central Manitoba (Duck Mtns.; Walley 1989a) and ne. Pennsylvania (Wyoming and Sullivan Cos.; Gross 1994). Adults feeding mobile young in nw. Oregon (Multomah Co.) in Jul 1976 suggests an isolated breeding event far from the normal range (Gilligan et al. 1994).
Abundant throughout most of its range, but local in mountains of the ne. U.S., where generally found above 600 m (Able and Noon 1976, Morse 1979a, Laughlin and Kibbe 1985). Also found in alder thickets and second growth, especially in northern and western parts of the range.
Winter Range
Generally found below 3,000 m in edges and second growth in n. South America east of the Andes (see Figure 2). Occurs from nw. Colombia, e. Ecuador, e. Peru, and n. Bolivia; east through Venezuela, Trinidad and Tobago, the Guianas (where scarce), and nw. Brazil (Amazonas, Roraima); to the mouth of the Amazon River (Sick 1993, Paynter 1995). Most common in the central portion of this range, and may occur along the northern coast primarily as a migrant (Bosque and Lentino 1987, Pashley 2010). Numerous sight records suggest that small numbers may also winter from se. coastal Brazil to ne. Argentina, and possibly se. Paraguay (Sick 1971, Paynter 1995, Greg Campbell pers. comm.). Small numbers recorded south of main range to 40°S in Chile and Argentina, the southernmost records for any migrant North American warbler (Dunn and Garrett 1997). Two January and one March record from Bermuda suggest overwintering there (Amos 1991).
Because the species migrates late, Dec records from e. U.S. are probably migrants rather than wintering birds, although there is a record of one visiting a feeder in Nova Scotia from 1 Jan to 4 Feb 1969 (Tufts 1986). Dec and early Jan sightings from the Greater Antilles and Trinidad and Tobago are also likely lingering fall migrants (Pashley and Martin 1988, McNair et al. 1996), although the possibility of occasional wintering should not be discounted.
Extralimital Records
Casual in fall (Sep–Oct) in Greenland (several records), Iceland (6), the British Isles (26), and islands off the coast of ne. France (2; Dubois and Homologation 1991, Curson et al. 1994).
Historical Changes to the Distribution
Completion of second-generation Breeding Bird Atlases in eastern portion of range allow comparisons of range between early 1980s and early 2000s. No significant changes were noted in New York (McGowan and Corwin 2008), Vermont (VCE unpubl. data), Massachusetts (Mass Audubon, unpubl. data), or Maritime Canada (BSC, unpubl. data), although in the latter atlas it appears the species may have colonized Prince Edward Island. In the White Mountains, New Hampshire, the lower elevational range has shifted down slope since 1993 despite an upslope migration of northern hardwoods and some low elevation birds (WVD). Although significant, the actual change in elevation was small and may be linked to increased basal area of balsam fir in the transition zone. The second Ontario Atlas detected a 44% increase in probability of detection (Cadman et al. 2007), primarily in the Hudson Bay Lowlands where the higher detection may be an artifact of higher survey effort. Extralimital breeding in Pennsylvania continued into the 2000s (Gross 2010), but has not been documented to date in Manitoba (BSC, unpubl. data).
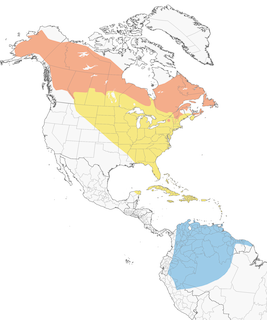
- Year-round
- Migration
- Breeding
- Non-Breeding
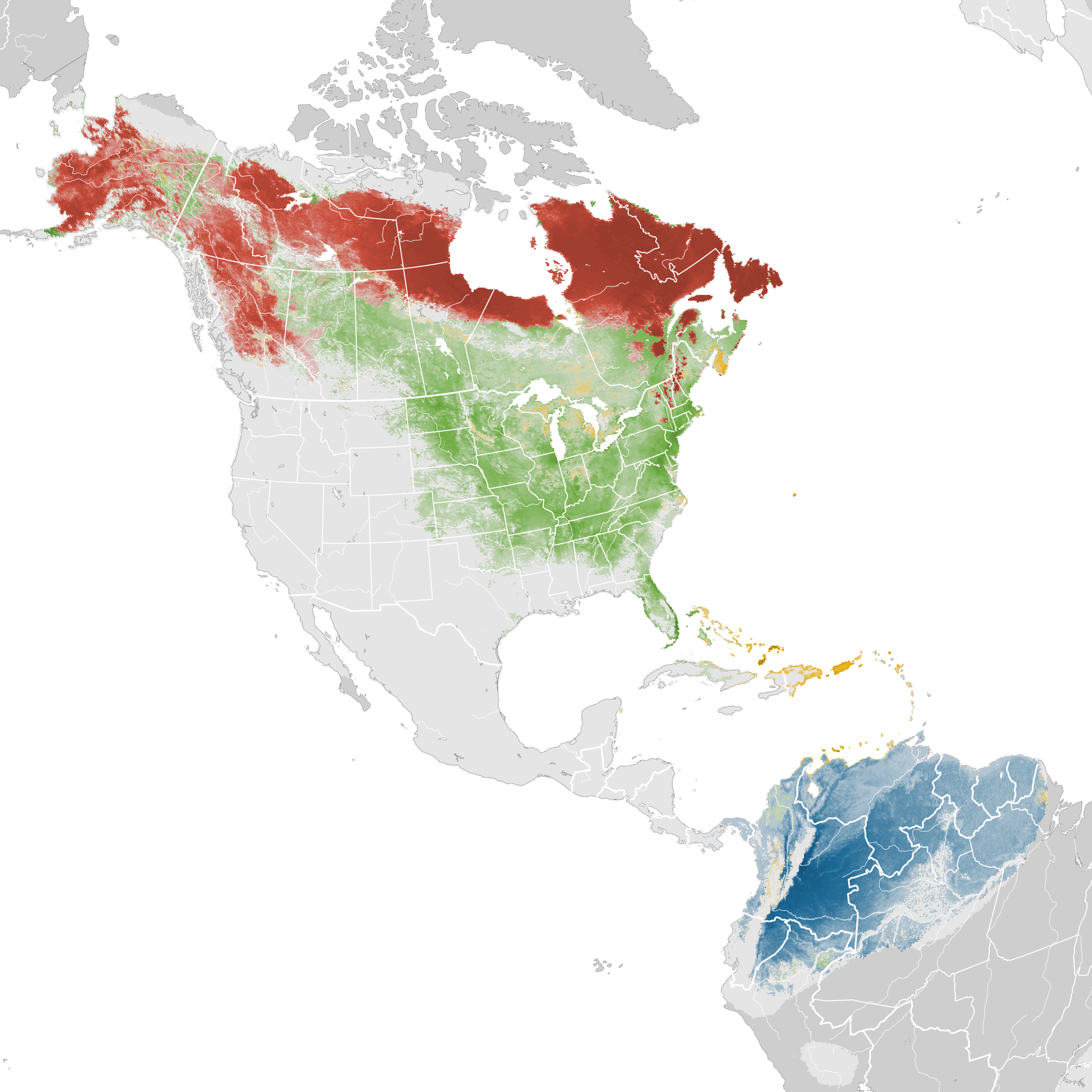
eBird Status and Trends

















